5.1��ǰ��
ά������ά����ʮ���������ܵ�һ��ͷ����л���������������ڲ��ܺϳɻ�ϳ������㣬����Ҫ����С����������ʳ�﹩����
ʵ�鶯����������Ȯ��������룩�Լ�����Ҳ��Ҫά���أ���˳���Ϊ������ά���ص�ʵ�鶯����۲�ȱ��֢��̽�������û�����
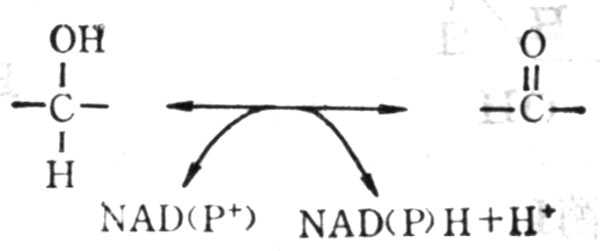
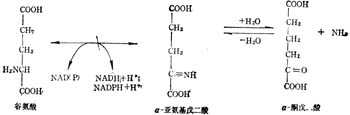
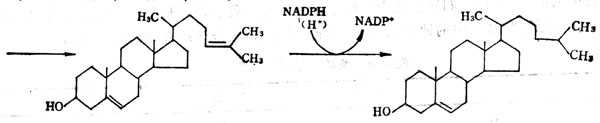
ά���ؿɷ�Ϊ֬��ʮ��ʮ��ˮ��ʮ��ʮ�����ࡣǰ����A��D��E��K��������ˮ��������֬����֬�ܼ��У���ʳ������֬�ͬ���ڣ��ڳ�������ʱ��֬������������ء���֬�����ղ�ʮ��ʱ�����������������к�����ǵ����մ�Ϊ���٣�����������ȱ��֢��ˮ��ʮ��ʮά����ΪB��ά���ؼ�ά����C��B��ά�����Ǹ�ø����ɲ��֡���B��ά���صĸ�ø���ڱ�5-1�С�B��ά�����е�B6�����ἰ��������ʳ���й㷺���ڣ�����ϸ���ֿɺϳɣ�����δ���ֵ��͵�ȱ��֢��
��5-1 ����B��ά���صĸ�ø
| ά���� | ��ø | ת�ƻ�ʮʮ��ʮʮ |
| ������� | ��ø��NAD+�� | ��ԭ�� |
| ͬ�� | ��ø��NADP+�� | ��ԭ�� |
| �˻��أ�ά����2�� | ���ص������ᣨFMN�� | ��ԭ�� |
| �˻��أ�ά����2�� | ���������ʶ���ij�ᣨFAD�� | ��ԭ�� |
| ���أ�ά����B1�� | ���������� ��TPP�� | ȩ�� |
| ���� | øA��HscOA��cOA) | ���� |
| �ܰ��أ�ά����B12�� | �ܰ��ظ�ø | ��� |
| ������ | ������أ���-N-��������-1-�����ᣩ | CO2 |
| ά����B6 | ��������ȩ | ���� |
| Ҷ�� | ����Ҷ�Ḩø�� | һ̼������ |
��ʮ��Ҫ��ʳ��������ά���ء�ʳ�ﲻ���ʳ�﹩Ӧ�����Ĺ��ң���Ⱥ������Ӫ��ȱ����������������ҳ������뵰���ʹ�Ӧ�����⣬ά����Ҳ����ȱ���������ǡ������������ͯ����ȱ��ά����A����ä�����ǰ���ҹ������ʮʮ��ʮʮ�ȣ�����ȱ����ʳ��ά����ȱ�������Ǻ��������鴨���Ϻ�����Ӫ�ĵ����з����������ά����ȱ���IJ�������ź�����������ƣ�ά����ȱ�����������١��������ҹ���ֲ��ʮ��ʮΪ��������ʮ��ʮʳ����٣�����ά����A���˻��ع�Ӧ��ƫ�͡���ˣ�����ɢ����ҹä�����ס�ʮ�ʮ��Ƥ��ά����A��B2ȱ����֢״��
�����˷���ά����ȱ������������ԭ��
��1��ά������ȡ������ ��ʳ���䲻����������ƫʳʮϰʮ������ά������ȡ���㡣���������ʳ�ﵥ������������Ϊ�������������ȱ�����Ƥ����
��2�����ղ�ʮ�� ���������ϵͳ�����Ļ��ߣ��糤�ڸ�к���������������ߡ�
��3������ϸ���������� ʹ��ɱ��ҩ���ʹ������ϸ���ܵ����ƣ��ϳ�ά���ص������٣�Ҳ������ijЩά���أ�K��B6������ᣩ��ȱ����
��4����Ҫ������ �����ڶ�ͯ����ʮ��Ͳ�ʮ��ʮ�ڵĸ�Ů���������Ͷ�������ֵĹ��˼��������Ⱥͻ���ʮ��ʮ����ʮ��ʮ�����IJ��˵ȣ���Ҫ����һ����Ҫ�ߡ�
��5��ʳ�ﴢ�漰����������� ���������ˮ����ʹˮ��ʮ��ʮά������ʧ�������?l��ʱ�Ӽά����B1���ƻ���ά����C�ڴ��漰���ʱ�����ƻ����ҹ���ʳ���߲˽϶࣬������ʳΪ��������ʵ����ȡ���Ȱ�������Ʒ�ļ���ֵҪС��
��Щά���ص�Ӫ������ָ�ꡢȱ��֢״�����δ�ʩ�Լ���ʮ��ʮ�Ƚ����й�ȱ�����ĸ���������������������ʮ��ʮӦ�������ڷ���ȱ�������ʹ�á�ʳ���ά���غ�������¼��ʳ��ɷֱ��ߣ��������롣δ�����ߣ����˶�Ҫ��̵Ľ��ܡ�
5.2������A
��1��ά����A ά����A�����ӻƴ���Retinol,VA��,����Ȼ���еĶ���ȫ��ʽ����������������������������������ܣ����ӻ�ȩ��Retinyl Aldehyde,VAȩ������Ӧ��Ч����¶���ӻƴ����ᣨMannosylRetiny Phosphate,MRP��Ҳ����ijЩ�������ܡ�VA������ķ���ʽ����ͼ5-1����ʮ��ʮ���ڱ�5-2��
ά����A������������������������MnO2��ʹ֮��Ϊȩ����ԭ��˫�ɽ�ά����A�ỹԭ��Ҫȩ���Լ���ȶ���������ȶ���ʹ�������˫���������С��ڹ���������£�ȫ��ʽ�ɱ�Ϊ���ȶ���11˳ʽ�칹�塣��ǿ�����γɶ���������塣
��2��ʮ��ʮ�ܲ��� ֲ����ʮ��ʮ�ܲ��������ڷֽ�Ϊά����A��������ά����A���������á�
��5-2 ά����A������������Ļ�ʮ��ʮ
| ��ѧ�仯 | ���� | �����ʮ��ʮ |
| ���� | ��Ȼ���˹���Ʒ | 100 |
| ���� | ȩ | 100 |
| ˳ʽ�칹 | ˳ʽ�칹�� | 23��75 |
| �� | �������� | 10��100 |
| ���� | ά����A2��3-�����ӻƴ��� | 30 |
| ʧȥ�� | Axerophene | 10 |
| ͪ | C21-ͪ | 3 |
| ȥ�� | ��ά����A | 3 |
| ����-CH2- | ������A | 1��5 |
| ��ˮ | ��ˮά����A | 0.4 |
| ���� | ������ | 0 |
| �⻯ | ����ά����A�� | 0 |
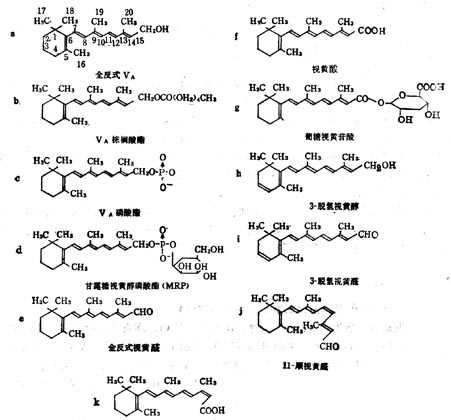
ͼ5-1 ά����A������ķ���ʽ
��-ʮ��ʮ���غ���������-������ͪ������-Ionone ring�����ĸ������ϩ��������ˮ����Ϊ������ά����A��ͼ5-2����ʵ������ʳ�Ц�-ʮ��ʮ�ܲ���ֻ��1/6�ұ�Ϊά����A����Ȼ�����л���ά����A��ͬ�����ǵ�ά����A��ʮ��ʮ���ڦ�-ʮ��ʮ�ܲ��أ��е�û����ά����A���ƵĻ���û�������ʮ��ʮ����5-3��5-4����
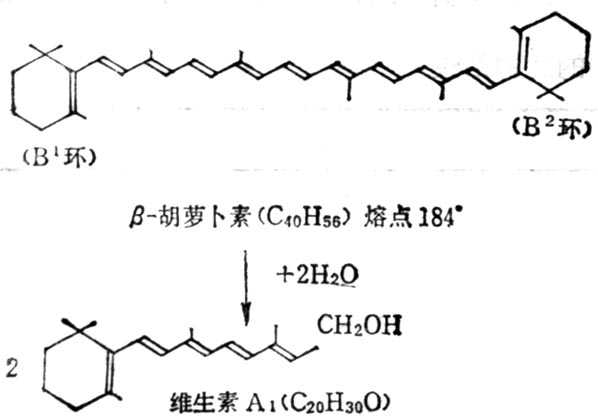
ͼ5-2 ��-ʮ��ʮ��ֹ��ˮ��Ϊά����A
5.2.2��ά����A��ʮ��ʮ�ܲ��صĴ�л
��1��ʮ��ʮ�ܲ��ص����� ʮ��ʮ�ܲ��ص�������Һ��С�������ա�����Һ����������ã���֬�������γ�������Һ���������ա����β����ٽ�ʮ��ʮ�ܲ�����������ϸ����������ϸ���������ϣ����ٽ�ʮ��ʮ�ܲ��صķֽ⡣ά����E���������������ɱ��������Ĺ���˫��ϵͳ����������ʮ��ʮ�ܲ��ؽ��뵽С��ϸ���ڣ��ڰ�����ʮ��ʮ�ܲ���˫����ø��Carotene Dioxygenase�������£���1mol�����뵽�м�λ�õ�˫���ϣ�����ֽ�Ϊ�ӻ�ȩ����Ҳ�ɴ�һ�˽���ֽ����ɾ�����ά����A��ͬ�Ļ��������ϳ���ȩ��apoʮ��ʮ�ܲ�ȩ�������ɵ�ȩ˫Ϊ����ø���û�ԭΪ����������С�����ζ���ʮ��ʮ�ܲ���˫����ø�����������С�����߽ϸߣ������ټƣ���Ϊ�ε�2������������Ϊ4��7��������ע���-ʮ��ʮ�ܲ��أ��ڸ���Ҳ��ת��Ϊ�ӻƴ���Ҳ���Ӵ�����ʽ���棬���������ޡ�
ʮ��ʮ�ܲ�����������ɢʮ��ʮ�ģ����������������С���෴��ϵ����ʮ��ʮ�ܲ��ض������ս���Ѫ���ڡ�Ѫ���е���ʮ��ʮ�ܲ��ؿɷ�Ϊ��-ʮ��ʮ�ܲ��ء�Ҷ���أ����Ѻ��ؼ�������ʮ��ʮ�ܲ��أ�����������������ɴ����ڸΡ�֬��������Ƥ����Ѫ������Ӳ���İ߿��С�
��5-3ʮ��ʮ�ܲ��ػ��ϳƵĻ��ṹ
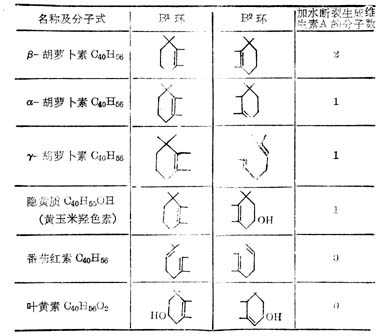
��5-4 ��ʮ��ʮ�ܲ��ػ�����������ʮ��ʮ���Ԧ�-ʮ��ʮ�ܲ������ʮ��ʮΪ100%��
| ��ʮ��ʮ�ܲ��� | ��ʮ��ʮ | ��ʮ��ʮ�ܲ��� | |
| ��-ʮ��ʮ�ܲ��� | 100 | ��-��ʮ��ʮ�ܲ��� | �� |
| ��-ʮ��ʮ�ܲ��� | 50��54 | ��-apo-8'-ʮ��ʮ�ܲ�ȩ | 72 |
| ��-ʮ��ʮ�ܲ��� | 42��50 | ��-apo-8'-ʮ��ʮ�ܲ��� | �� |
| ��-��ʮ��ʮ�ܲ��� | 20��40 | ��ˮҶ���� | 21 |
| 3��4ȥ��-��-ʮ��ʮ�ܲ��� | 75 | �����أ������ʣ� | 57 |
| ��-ʮ��ʮ�ܲ��� | 21 | ��-˳�칹���ȫ��ʽ�Ļ�ʮ��ʮҪС | |
| ��-ʮ��ʮ�ܲ��� | 50 |
��2��ά����A�����ռ�����ά����AΪ�������գ���Ҫ���������ʱ�ʮ��ʮ�ܲ���Ҫ��7��30����ʳ���е�ά����AΪ��ʽ����������Һ����ʮëʮˢ״Ե�е��ӻ���ˮ��ø�ֽ�Ϊ����ʽ���뵽С��ʮ���ڣ���Ϊ����ϸ�������е���ø���������ϳ�ά����A�����������ȡά����A3��5h�����մﵽ��ʮ�塣ά����AҲ��Ҫ���Ρ�ά����EҲ�ɷ�ֹά����A�����ƻ���ά����A��ʮ��ʮ�����������ܰ�ϵͳ���͵��Σ���ʽˮ�����Σ�Ȼ����������Ϊ�����������ʵ��ϸ��������ȡ���½����ά����A��л��һ����ά����A��ʵ��ϸ��ת����֬����ϸ����Lipocyte��,����ϸ������������֬�Σ���ά����A��ȡ������Щ��֬�δ���ࡣ85%��ά����A����֬���У�����һЩ�ڸ߶������С�
ά����A�������봢�������ϴ���ÿ������20��000IU����ά����A�����ٶȺܿ죬VA����֬���Ľ���Ѵﱥ�ϣ�����������ʽ���뵽Ѫ���С�ÿ�ո�����50��000IU��4�ܺ������ά����A��ʳ3�ܣ�Ѫ����ά����A���������߶࣬��ά����Aֵ������һ����������֯����Ҫ��ά����A�Ӹ����ͷž�ʮѪ��������֯����һ�����ʣ��ά����AҲ�Ӹε�Ѫ���С�
�ߵ�����ʳ��������ά����A�����ã��������ά����A����Ŀսߣ�Depletion����������Ӫ����ʮ��ʱ��ά����A�����ռ�ʮ��ʮ�ܲ���ת��Ϊά����A���������ܵ�Ӱ�죬���ʹά����A�ĸδ��������͡����п�ʹ���������ӣ���������17��21��ʱ���δ��������2����Ϊ����ά����A�ḻ�ij�ʮ��ʮ��ʮ��ʮ֭������̥��ά����A���������٣�����ĸ��Ӫ�������Ӱ�졣����Ӥ����ά����AŨ�Ƚ�Ϊ��ʮ��1/10��60���Ժ����ֵ��ڳ�ʮ�ˡ��δ�����������ʮ��ʮ���й�ϵ�����������϶ࡣ��ά����Aȱ�����ϣ�����δ���ս߱ȴ���Ҫ�졣��ʮ��ʮ��Ѫ��ά����Aˮƽ��Ůʮ��ʮ�Ըߡ�ҹä�ķ���������ʮ��ʮ�϶ࡣ������Ҳ�ܴ���ά����A����������Ϊ�ε�1%����ɫ����Ƥ��֯�ڵ�ά����A������ʽ���ڵģ�רΪ����Ĥʹ�ö���������ս����ʱȸ�����Ҫ��һЩ��
��3��ά����A�����䵱����֯��Ҫά����Aʱ��ά����A�Ӹ����ͷų��������䵽����֯������������Ƚ����ڴ���ά����A������øˮ��Ϊ��ʽ�����ӻƴ���ϵ��ף�Retionl BindingProetin,RBP����ϣ�����ǰ���ף�Prealbumin,PA����ϣ��γ�ά����A-RBP-PA����ʮ����뿪���ࡣ��ʮѪ�������֯��ά����A��һ������±����뵰���ʽ�ϣ�ʹ֮����ˮ��ʮ��ʮ�������ȶ������⣬���ɼ���ά����A��ϸ���Ķ�ʮ��ʮ����ϸ��Ĥ�Ը���ʮ����ʶ������������δ��ϵ�ά����A��ʶ���������¶�ά����A����ȡʧȥ���ƣ�����ά����A����ϸ����������ʮ��ʮ��
RBPΪ��ʵ��ϸ���ϳɵĶ��ģ�������21��000����Ӿ�ڦ����֡���ÿ���ܺϳ�190mg��һ����RBP��һ����ά����A��ϡ�ά����A-RBP-PA����Ѫ���У����ڷ��ӽϴ��ܴ������˳���Ѫ����RBP��ˮƽ��ά����A��Ӱ�죬�����ά����A��������ʱ��RBP������ڸ��ڡ�����Ѫ��RBPˮƽΪ40��50��g��m-1��ά����Aȱ����ҹä֢״����ʱ��RBP��Ϊ20��g��m-1��������ȱ����RBP�ϳɼ��٣�ά����A-RBP�����γɣ�ά����A��ϵ�RBP-PA������Ϊ����ֵ��20%�����ѱ�ʮ��ʮ����������ά����A��ϡ�PA��Ѫ��Ũ����ά����AӪ��״����Ӱ����٣�������ֵΪ200��300��g��m-1����������RBP��ϣ�����Ҳ���״���ؽ�ϣ�������Ӱ�졣
ά����A-RBP-PA����ʮ����Ѫ������ճʮĤ����ʮ�ס���Ĥ����Ƥ��֯�Ȱ�ϸ����ϸ��Ĥ����RBP��������ȫ������RBP��ϣ�����ά����A�ͷų���������ϸ���ڡ�RBP��ά����A�ֿ����ѱ�ʮ��ʮ��ɥʧ��ά����A��PA��ϸ��Ĥ������Ľ�����������������RBP����С���п��˹���������С�������գ�Ϊ��Ƥ��ϸ������ȡ����������ø���зֽ�Ϊ�����ᡣѪ���л���һ��δ��PA��ϵ�ά����A-RBP������ԼΪ1��g��m-1���ⲿ�ֿɴ���С���˹�������С���������գ�����ά����A�ͷų������������������ά����A���ۣ�Ȼ�������������Ρ�
ά����A���뵽��ϸ����������ϸ���Ӵ���ϵ��ף�Cellular retinol Binding Protein,CRBP�����ϡ�ά����A������������в���Ҫ��RBP���ϣ�������ϸ���ڱ����ӻ����ϵ������ϡ�
�β����ߵ�Ѫ�����ϳ��ܵ�Ӱ�죬RBP��PAˮƽƫ�Ͷ�Ӱ��ά����A��Ѫ��ˮƽ����������RBP�ֽ��л������ʹRBPֵ���ߡ�������ά��ʮ��ʮ���ߣ�����ά����A�������ʵ�����Ѫ����RBP��PA��ά����A��ˮƽ���͡�RBPҲ�����ڵĵ��ڣ���ʮ��ʮ����Ѫ����RBPˮƽ�ȴ�ʮ��ʮ�ߡ���ʮ��ʮ�ڻ�ڷ�����ҩ�ĸ�Ů��RBP���б仯����������RBPֻ�г�ʮ�˵�һ�롣����ˮƽһֱά�����ഺ�ڣ��Ժ����ߡ�
��4��ά����A�ֽ��л����ʮ��ά����A�ֽ��л;��������ͼ5-3���ӻƴ�ͨ������ת��Ϊά����A�ᣬ����һ�����칹Ϊ��˳ʽ��ȫ��ʽ���˳ʽ�߾�Ϊά����A�Ĵ�л�����ά����Ƥ��֯�ֻ��Ļ�ʮ��ʮ�������ڲ��ܴ��棬�ܿ���ʧ��ע��ά����A��4h����ֻʣ��10%��24h��������ޡ��Ժ�5��6λ�������γ�5��6����ά����A�ᣨ5��6-epoxyretinoic acid������4λ�ǻ������ٽ�һ�������γ�4-��-��ά����A�ᣨ4-oxo-Retinoicaxcid�����γ������ӻ����ᣨretinoylglucronide��������ע����ά����A�������֭��60%Ϊά����A�ἰ�����������ᣬ����40%��Ϊ13˳ʽ�����ӻ����ᣬ��������5��6λ�ӻ���ά����A�ἰ���������ӻ�����Ϊ0.2~0.4��g����Щά����A��л�ӵ�֭����С�����γɸγ�ѭ������������ע����ά����A��2h����С���еķ���ʮ��ʮԼռ������7.2%������������߽�Ϊ2.4%����������Ҳ��ά����A��л�����θ���Ա�ǵ��ӻƴ����������������г��������⣬����ȫ��ʽά����A�ἰ5��6λ����ά����A�ᣬ����Ѫ���в�δ���ֺ��ߣ������������ÿ��ܽ��ڰ���֯���������з��������Ļ�ʮ��ʮ��Ϊά����A��֮1%����������ں̡ܶ�
ͼ5-3 ά����A��������л�Ŀ���;��
ͼ5-4 ά����A��������л�Ŀ���;��
��������ά����A��㼰���ж������л�����ʮ����2/3��ʮ����������ϩ���������ı䡣Ҳ��15-C����ΪCO2���еIJ��������ֽ��CO2��������Ʒ���������20��gά����A�������ʮ��������������Ϊ����Ϊ��Ҫ��ʮ��;���������ʮ�����仯�ϴ�2��3��ﵽ��ʮ�壬ԼΪ������9%��Ȼ���½�����С������������ʮ��24h��Ϊ������8.7%������һ��Ϊˮ��ʮ��ʮ�ߣ�һ��Ϊ֬��ʮ��ʮ�ߣ�����ʮ�������½����ڰ���ﵽ�ȶ���ԼΪ0.55%�����д�л�����-������ͪ�����ֿ�������������ʮȥ�����е�˫���ɱ��ϣ���Ҳ������ľ�̡�ˮ��ʮ��ʮ��л��Ŀǰ�˽���١�
���ڴ���ļ������ά����A���ܱ����䵽��Ҫά����A�İ���֯��ȥ����ˣ�����ά����A�İ���֯�С���ˣ�����ά����A�Ĵ�������Ӱ��ά����A�Ĵ�л�ʣ���ά����Aת��Ϊ�����ά����A��л��֮������SauberLich�ⶨ�˵Ĵ�л��ÿ��570��1250��g�����ֲ������ڸδ�������ͬ���¡�ע����ά����A���������У�����ά����A�����ֲ�ͬ���ʴ�������ʮ�����༴�����ֲ�ͬ��л�⣺��л��1���������յ�ά����A����л��2��������ʮ��ʮ�ͷǹ���ʮ��ʮ��ά����A����л��3��������ʮ��ʮ�Ĵ�л���ڸ��ڴ���սߺ�Ĵ�л�ʣ��Ŵ�������֯�Ĺ���ʮ��ʮ��л�ʡ������յ�ά����A�����ڴ����߲�ͬ��������24h�ڲ���ϣ�Ѫ������������ռ90%����Դ�߽�ռ10%��������ϣ��������ڰ������ȫ��ϡ��������ά����A�������٣�����������Ѫ���б�������ʱ��ϳ�������δ�����Ϊ0.6��g��m-1ʱ������������������ά����A�Ѿ��սߣ���Ҫά����A���䣬���ܴ����ڸ��ڣ�������������֯�����ȡ���ά����A�������϶�ʱ���δ�����Ϊ18��g��m-1��60%���ڸ��ڡ��Ӵ�л��2�������ǹ���ʮ��ʮ�Ĵ�л���������δ���ά����A�ķֽ⡣����ά����A�սߵ�������ά����A��������ָ�����仯�������ά����A�δ�������Ϊȫ��������Ҫ������������Ҳ�ɿսߣ���ʮ��Ҳ����ˣ��������ߴ�л����ߣ��������ߣ���л��Ҳ�ߡ�
5.2.3����������
ά����A����ά��������������ʮֳ���Ӿ�������Ⱦ�Ĺ��ܣ��������û���������δ��ȫ�����
��1��ά����A���Ӿ��Ĺ�ϵ����Ĥ����������ϸ�������Ӹ�ϸ������ϸ��������ǰ�������࣬�밵��ʮʮ��ʮʮ�йأ����������٣������Ӿ���ɫ���йأ����Ƕ�����ɫ�أ����ӵ�������ɫʮʮ��ʮʮ��ɡ����ֶ����ϸ�����ӵ��ײ�ͬ����ɫʮʮ��ʮʮ�ɲ�ͬ���͵�ά����Aȩ��ɡ��Ӹ�ϸ����κ�����ɫ���Ǹйⲿ�֡��Ӹ�ϸ��Ĥ�������ϵ����ݣ������γ�Ƭ��Ĥ��ɣ�ÿһ��Ĥ��������֬�����Ĥ������֬�����Ĥ���ɳ�Ϊ˫����Ĥ����ɫ����Ƕ������֬��˫����Ĥ�С���ɫ�����Ϻ��ʣ���ά����Aȩ���ӵ���϶��ɡ�ά����Aȩ��ά����A�������������칹ø����ʹ���Ϊ11-˳ʽά����Aȩ��ά����Aȩ��ά����A�������������칹ø����ʹ���Ϊ11-˳ʽά����Aȩ��ά����Aȩ���ӵ���֮���һ���ṹ��11-˳ά����Aȩ��ȩ�����ӵ�����������ΰ����γ�ϣ��Schiff base��������֮�����ӵ����ṹ�ı䣬����������Ķ����ṹ���ںڰ��зdz��ȶ���������ʱ��һ�����Ϻ��ʽ���һ�����Ӻ�ά����Aȩ��11-C��Ť��ת��ȫ��ά����Aȩ���ӵ������幹��Ҳ�����仯��ά����Aȩ�־���ϸ����˵�ά����A��ԭøʹ�����Ϊά����A��Ȼ����ɫ����Ƥϸ����������ø����������������ɫ����ϸ���ڣ���Ҫʱ���칹Ϊ11-˳ά����A������Ӧ�������෴��ʽ���У����γ����Ϻ��ʡ���ʱNa+���Ӹ�ϸ���ڶ��Ƶ���Σ��γɰ�������Dark current flow���������Ϻ��ʾ����칹���Ϊǰ�����Ϻ��ʣ�Prelumirhodopsin��,����ϵ�Ca2+���Ӹ�ϸ������ͷų�������ֹ��Na+������������������������ѹ�IJ��������ֵ�λ��ʹ�̼���ǿ�������з�����У�Ҳ��������Ĥ��ͼ�ⶨ��
ά����A-RBPͨ������ĤѪ�ֲܷ�������Ĥ��Ƥ��֯����ϸ��������ά����A-RBP���ܵ㣬��ά����A�����ǿ����RBP�������ά����A���뵽ɫ����Ƥϸ��������ʽ�����ڰ������͵��ڡ�����Ӧ�����У���һ���ά����AΪ11-˳ʽ�����Ӹ�ϸ�������������������ά����A������60%Ϊ11-˳ʽ�������Ϻ���Ҫ������Ĥϸ������CRBP��������Ĥ����ȱ�����ֵ��ף����ά����A���ܽ��뵽ϸ���ڡ�
ι��ά����Aȱ����ʳʱ�����ȸ��ڴ����ά����A�ͷŹ���������֯���εĴ������½���Ѫ��ά����A���������δ����������ս�ʱ��Ѫ��ά����Aˮƽ���½�������������Ĥ��ɫ����Ƥ��֯��ά����A���ۣ����Ϻ��ʵ�ά����Aȩ�½�������ɫ����Ƥϸ����ά����Aȱ��ʱ������ҹä֢���֡����Ӹ�ϸ�������Ϻ��ʼ��٣�Ӱ�������գ�����Ҳ���١������Ϻ���Ϊ����ֵ֮74%����������14������Ϊ����ֵ֮16%����������680����ά����A�dz�ȱ��ʱ���⵰��Ҳ���٣���ϸ�������ܽ⡣
��2�����ǵ����ʺϳɵ�Ӱ�� ȱ��ά����A�Ķ����ij��֯����С������Ĥ��������Ƥ��֯��Ѫ��ȵ������ǵ����٣�����ά����A��ά����A����Դٽ���ϳɡ�ϸ��Ĥ����ĵ�����ҪΪ�ǵ��ף�ϸ��Ĥ�Ĺ�����Ӵ����ơ��ֻ���ʶ�����������������ǵ����йء�Ĥ�����ǻ��ӻƴ����ᣬ���ԣ�ά����A������Ĥ���ǵ��ϳ��йء�������Ҳ���ǵ��ף�ά����AӪ��״��Ӱ�����߹��ܣ���������йء�
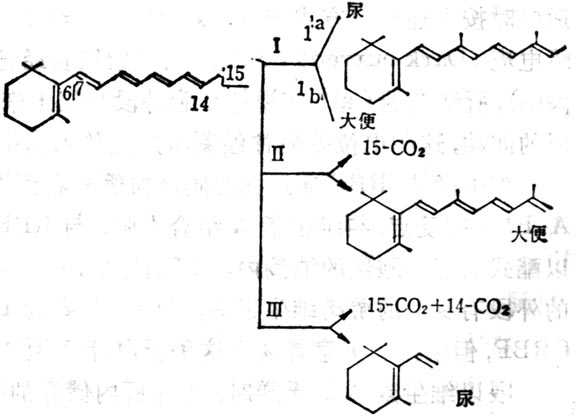
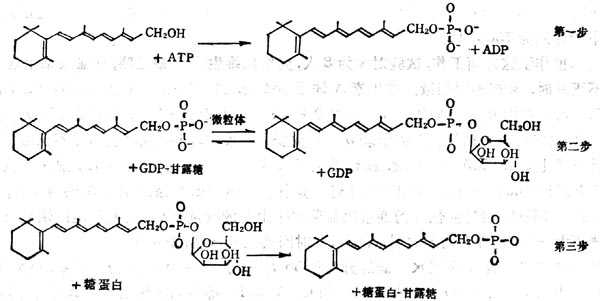
�ǵ��ĺϳɲ��裬����Ϊ�ϳɶ�������Ȼ�����뵥�ǣ�������������ϵ�˿���հ�����ǻ����Ŷ�������ż���γ��ǵ��ס��ǵ����е���Ϊ��¶�ǡ������ǡ���ʮ��ʮ�ǡ������Ǽ��������Ѱ����ǵ��ĺϳ���Ҫ֬��-����Ϊ�м��壬����֬���Ϊ�����ϩ����Ķ��ƴ����ӻƴ��������Ǵ�GDP-���ǻ�UDP-����ת�Ƶ�ά����A����ƴ��ϡ�������Ϊ��¶�ǽ�ϵ�ά����A�ϣ������ɸ�¶���ӻƴ�������(Mannosylretinylphosphate,MRP)������ϵ����ƴ������ɸ�¶�Ƕ��ƴ���������Dilichylphosphomannose,DMP�����ٽ�����ת�����ǵ����ϡ�
ά����A���ǵ��ϳ��б仯��Ϊ�������裺���γ�ά����A��������Retinylphosphate,RP��������RP�ϳ�MRP����һ�����������ںϳɣ��۽�����ת�����ǵ����ϣ�ͼ5-5����
ͼ5-5 VA���ǵ��ϳ��е�����
5.2.4����Դ
����ĸΡ���������ʮ��ʮ�еĶ���ά����A�����Ը�����ḻ����ɫ�߲˼����ɫ�߲���ˮ������ʮ��ʮ�ܲ��أ�������ʮ��ʮ�ܲ��ص�����Ч�ò�һ�����Ԧ�-ʮ��ʮ�ܲ�����ߡ���Ӧ��Ϊ1500��4000IUʱ��ʳ����1��g��-ʮ��ʮ�ܲ����൱��0.167��g��ά����A������ʮ��ʮ�ܲ��ؽ�Ϊ��һ�롣ֲ������Щ�ƺ�ɫ�ز���ά����A��Ч�ã���5-4����
ʮ��ʮ�ܲ���Ϊ֬��ʮ��ʮ�ģ��ڶ�ֲ������֬�����ϣ����������п���ø�⣬�����ʮ��ʮ�ܲ��رȽ��ȶ����ҹ��ij��˷�����ʮ��ʮ�ܲ��صı�����Ϊ76��94%�������ʱ��̣���ˮ���٣�ʮ��ʮ�ȵͶ��ɼ�����ʧ��ֲ���ϻ������У�ʮ��ʮ�ܲ���ת��Ϊ���������Ϊ�����ƻ��ĵ�һ�����ӹ�������������Ҳ�����칹�����������ʮ��ʮ��ʳ��ӹ���ʹ�������ᣬ������ʮ�ɻ�����ʮ��ʮ�ܲ���Ҳ���ƻ����á�ʳƷ��������ҲӰ��ʮ��ʮ�ܲ��ص����á�ʮ��ʮ�ܲ�Ƭ��ʮ��ʮ�ܲ���76%�ɴ����ʮ�������Ƚ�����ֻ��44%��ʮ����
5.2.5����Ҫ�����о�
VA��ȥ�Թ��ʵ�λIU��ʾ���������ӻƴ�����RE��ʾ��
1�ӻƴ�����=1��gά����A
���δ�սʱ��Ӣ��ҽѧ�о����16����ά����Aȱ����ʳ��Ϊ�����飬Ϊ��61/2��25���¡�7��Ϊ�����飨����2��ÿ�ղ���25IUά����A��5��ÿ�ղ���5000IUʮ��ʮ�ܲ��أ�������������ȱ��֢״���䲻ͬ������ά����A��ʮ��ʮ�ܲ����Բⶨ��Ҫ�����ڴ��о��й۲�ָ��϶࣬����Ѫ��ά����A������Ӧ��Ϊ��������������Ҫ��Ϊ1300IUά����A��1500IUʮ��ʮ�ܲ��أ��Ӵ������ʮ���߲��������ڣ���������Ҫ��Ϊ2500IUά����A��ʮ��ʮ�ܲ���Ϊ3000IU��������ʳ���ʮ��ʮ�ܲ�����Ч������һ��������������Ҫ����ʳ����ʮ��ʮ�ܲ��ؼ�ӦΪ7500IU��
����Ҳ�����ⷽ�湤�����������Ϊ8�ˣ��ȸ���ά����Aȱ���ţ���Ѫ��ά����A������������ʱ�����䲻ͬ������ά����Aȱ��184�죬Ѫ��ά����A��57��78��g%����20��32��g%��������Ѫ��ά����A�Գ�ȱ��֢�����ʱ��Ϊ361�죬�Ϊ771�졣Ѫ��ά����AŨ����ȱ��֢�Ĺ�ϵ���ڱ�5-5��Ƥ���ǻ��������磬����Ĥ��ͼ���������ֽ���������ά����A37.5��150��Ѫ��ά����A���������ԣ�����300��g��Ѫ��ά����A��������������600��g�ߣ���������������ʮ��ʮ�ܲ���1200��g��Ѫ��ά����A��������������2400��g�����Ͽ죬����Ӧ�ָ��ļ�������ˮƽ�����ļ���ҪС������Ĥ��ͼ�ָ������ļ���Ҫ����һЩ��������ͬ�ȵļ�������ָ�ʱ��Ҫ�ӳ�һЩ����5-6����
�ҹ�����ά����AѪ��ˮƽ�͵�������56�ˣ�Ѫ��30��65IU�����ֳ�7�飬ÿ��8�ˣ��ֱ����400��2000��3100��3500��3800��4500��5500IU�Ȳ�ͬ����ά����A��������Ϊ11�ܣ��۲�Ѫ��ά����A������Ӧ������ä��ı仯��ÿ������2000��3000IU�ߣ�Ѫ��ά����Aˮƽ������������δ�ﵽ100��120IU%��3500��5000IU�ߣ���9��ʱʹѪ��ά����A�ﵽ100IU%��11��ʱά����107��130IU%���������δ�۲쵽����Ӧ�ı仯������������ڣ����������������������ä�������������ȡ��Ϊ400IU�ߣ�����ä���������һ��1��000��000IU�������24h�ڻָ���������ȡ��2000��3000IU���飬��11��ʱ�ָ���������ȡ��3500IU���ϵĸ��飬��5��9��ʱ�ָ���������ˣ�ά����A��Ӧ��Ϊ3500IUʱ��������ʹѪ��ά����A�ﵽ��������������ä��Ҳ����������������ʮ������ĵ��������֡�������ͬ������ά����A������Ҫ����Ϊ1200��1300��g��
��5-5 ����ά����Aȱ��֢״����Ӧ��Ѫ��Ũ��
| ά����AѪ��Ũ�Ȧ�g% | |||
| ������ | ����Ӧ������ | ����Ĥ��ͼ������ | Ƥ���ǻ� |
| 1 | 22��24 | 9��11 | 29��35 |
| 2 | 25��30 | ��� | 32��37 |
| 3 | 20��25 | ��� | 29��38 |
| 4 | 4��5 | 7��10 | 20��25 |
| 5 | 4��8 | 4��8 | 5��10 |
| 65 | 30��35 | ��� | 25��30 |
| 7 | 2��5 | 5��9 | 13~ 20 |
| 8 | 3 | 5��10 | 20 ~ 24 |
��5��6 ����ά����A��Ҫ��
| ά����A��Ҫ�� | ||
| ά����A����g/�գ� | ��-ʮ��ʮ�ܲ��أ���g/�գ� | |
| ����Ӧ���� | 150��300 | 300��600 |
| ����Ĥ��ͼ���� | 300��600 | 600��1200 |
| Ƥ������ | 600 | 1200 |
| Ѫ��ά����Aˮƽ����20��g% | 600 | 1200 |
| ����30��g% | 1200 | 2400 |
��Ѫ��30��g%���ϸδ�����Ϊ�е�ˮƽ��ά����A��л��Ϊ570��1250��g��
5.2.6����ʮ��ʮӦ��
ά����A�����Է���ȱ�����⣬����������Ƥ���ǻ�������ijЩ����Ԥ�����á�
��1��ά����A��Ƥ����ά����A������Ƥ���ǻ���ά����A��Ҳ�����ֹ��ܣ������������ڸ��ڣ����䲻��ҪRBP���ɽ�����֯�У�Ѹ�ٴ�л���ܿ��������ʧ����ʮ��ʮ���١�������ά����A�ἰ������������ʮëʮ�ҽǻ�����ȡ���-˳ʽά����A����ʮ����ά����A���������Etretinate����Ч�ֱ�Ϊȫ��ʽά����A���2.5��10������-˳ʽά����A����������Ƥ֬�γ��������Ч����Etretinate����ţƤѢ�������������Ѹ�ٴﵽ��Ƥ��Ƥ��
��2��ά����A��ά����A��ά��������Ƥ��֯�ķֻ���ά����Aȱ����ijЩ��֯����̬�仯��ijЩ��Ƥ��֯��֢�����ڲ��䣨��ǰ�ڣ������������Ƥ��֯��״���Ρ���ˣ�ά����A��������һ���У���ϸ�����ת�����á�ά����Aȱ������������ijЩ����֯���°����ʵ���ʮ��ʮ��ʮ��ά����Aֻ�ڰ�ǰ�������ã��������ڰ����ֺ�����Ч�����������֤ʵ����˵����
��ȡ������ά����A�������ڸΣ�ѪҺ�к����ܵ�RBP�� ���ƣ�������֮֯�����ޡ�Ԥ������Ҳ����ά����A�ᡣ��������������ʮ��ʮ��С�����������ڰ���֯��ά����A��������٦�-������ͪ���ĸı䣬�±�Ϊ�������Ϊ����������ϩ������3������������ϩ���ἴEtretinate��������Ч�ؼ��������ߵ�������֯���Σ���ʮ�װ�������Ҳ���ͣ��ڲ����ϵĸı䣬9��11��13����˫��Ϊά����A��ʮ��ʮ��Ҫ���֣����뱣����˳ʽ�칹���л�ʮ��ʮ����-˳ʽά����A��������ʮ��ʮӦ���ڷ��ΰ�ʮ�װ����������������Ķ�ʮ��ʮ��ά����A������ά����A�һ�����Ҫ��ĩ�˼�ʮ��ʮ��ʮʮ��ʮʮ�ĸı䣬ά����A����������ʮ��ʮ�ͣ���ʮ��ʮ�ߣ����л����٣�����ʮ��ʮ���н϶࣬������Ԥ��ʮ��ʮ�ٰ����յ�������δ������ʮ�塣
ά����A������ϵͳҲ��Ӱ�죬�������Լ����ֿ����йء����Ҷ�һ��������İ�Ҳ��Ԥ�����á�С�������ϸ����ֲ��С��δ����ά����A�ߣ�75%��3��5���ڷ�չ������������ά����A��ÿ��5000IU��������5���߽�5%�����������ⷽ�湤��Ҳ�ڽ��С�
ά����A��Ԥ�����ÿ����������������ʹ����ı䡣ά����A��ϸ������ά����A��ά����A���ϵ��ף�CRBP��CRABP��,ά����A��ά����A����֮��Ϻ��ܱ�Я��������ϸ���˴Ӷ��ı����Ӱ��ϸ���ֻ���CRBP�������Ρ�С�����Ρ�Ƣ���ۼ�ʮغʮ���н϶࣬��Ѫ�塢�ԡ����⡢֬���������н����١�CRABP����̥��δ�ֻ�����֯�ж࣬��Щ����֯�У�CRABP������������֯����Ҫ�ࡣ��֯��CRBP��CRABP֮������ά����A��ά����A�ἰ����������������á���ͬ����֯��ά����A����������ʮ��ʮ��ʮ����ȡ����ϸ����CRABP֮��ͬ��
ά����A��ʮ��ʮ�ܲ���Ҳ�����ڸ������ơ������г������Ƽ����ƺ�ʹ��ϸ��ˮƽ������ˮƽ��ά����A��ʮ��ʮ�ܲ��ضԲ���İ�ϸ���ķ������������ã��Ӷ����ٸ����ʻ��ӳ�DZ���ڡ�
��3��ʮ��ʮ�ܲ����방 ���в�ѧ�����з����ΰ��ķ�������ʮ��ʮ�ܲ�����ȡ�����෴��ϵ������θ����ǰʮ��ʮ�ٰ�Ҳ�����ƵĽ����������ʡ����66�����ϵ�������5���ڵ������ʣ�����ʮ��ʮ�ܲ�����ȡ�����ߣ�����Σ��ʮ��ʮ��Щ��
��ǻճʮĤϸ�����̡��ơ�X���估�����Ƶȴ̼��£�������ϸ����micronuclei cell��,����ϸ����DNA����Ⱦɫ������ı�ǣ����ܷ�չΪ��ǰ�ڵIJ��䣬�������ά����A��ʮ��ʮ�ܲ������ƾ��ߣ����Լ�������ϸ���İٷ��ʣ������ʱ�����������ٵ����ࡣ
��С�������У��������-ʮ��ʮ�ܲ��ؿ�����2�������յ���Ƥ���°����ã���Щ��ʮ��ʮ�ܲ��������á�
һ����Ϊ��ʮ��ʮ�ܲ�����С��������ת��Ϊά����A�����н��������Ŀ�ת�䡣�����С���������Ԧ�-ʮ��ʮ�ܲ��غμ�Ƥ���пɻ���ά����A�������п��ܦ�-ʮ��ʮ�ܲ�����ת��Ϊά����A��������á���Щ����Ϊʮ��ʮ�ܲ�����Ϩ����̬����Singlet O2������ʮ�ɻ������ֵ���̬���ķ�Ӧʮ��ʮǿ���Ի����и����ã���ֲ���л�в�����ֲ������ʮ��ʮ�ܲ�����Ϩ���������²����������к����ʡ�ʮ��ʮ�ܲ��صĿ������ÿ���������Щ���á�
ʮ��ʮ�ܲ��ش�����ȡ����Ƥ��֬������ʹ���ʻ�ɫ�⣬��δ������������ʮ��ʮ��
5.3������D
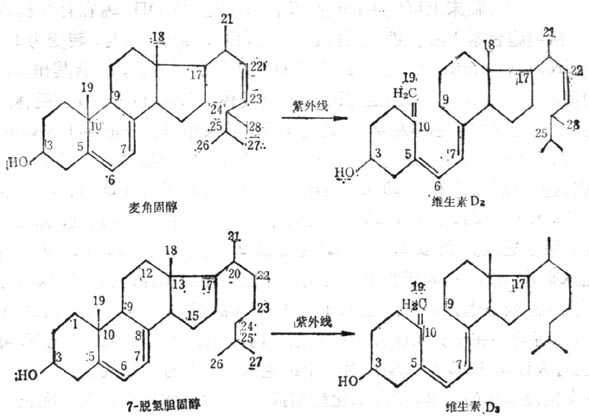
5.3.1���ṹ��ʮ��ʮ��
ά����D��VD���ǻ�����������������ά����Dԭ��ProvitaminD����������270~300nm�����γɡ�����Ƥ��7-����̴�����ĸϸ���е���ǹ̴�����ά����Dԭ������������ֱ�ת��Ϊά����D3��ά����D2���٣����˹������߶�Ϊ���ͣ�ͼ5-6����ά����D��������շ�Ϊ265nm���Ƚ��ȶ����ܽ����л���ý�У�������ٽ��칹���ã�Ӧ�����ڵ���������������价���У�����Һ�ӿ����������ȶ���ˮ��Һ�������ܽ�������ȶ���˫��ϵͳ��ԭҲ����ʧ������Ч�á�
ͼ5-6 ���̴���7-��������̴���������������ת��Ϊά����D2��ά����D3
5.3.2����л
��ʳ���е�����ά����D����֬��һ�����գ����ղ�λ��Ҫ�ڿճ���س�����֭���������ա�֬�������ܸ���ʱ������ʮ��ʮ�����ס�֬������������������Ӱ���������ա����յ�ά����D��ʮ��ʮ�������ϣ����ܰ�ϵͳ���䣬��Ҳ����ά����D���䵰�ף���-�ײ��֣�������Ѫ�������䡣��Щ���-֬�������ϣ��ڷ�ά����D��ʮ��ʮ������ϣ��ȴ�Ƥ���������뵰��������ڷֽ⡣
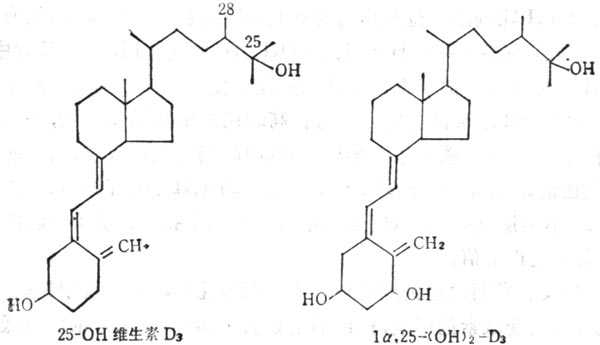
��ά����D�˵������У��������о�����øϵͳ���ã�����25λ�ǻ����γ�25��OH��D(25-hydroxy vitamin D3)(ͼ5-7)�������������֯Ҳ����ȡά����D��25-��OH��D3�������֯��ά����D��25��OH��D3����������Ѫ���ж࣬�������֯��Ҫ���ɽ����ͷų�����������֬����֯����࣬�ͷ��ٶ������������ؼ��ᣬ֬������ʱ������Ҳ���ͷų���������ע��ά����D���Ͽ����Ѫ�����뵽��֯�С�Ѫ����25��OH��D3��ע���1��3��ﵽ��ʮ�壬��Ũ�ȿɴﵽ20��40ng��ml-1,��߿ɴ�80ng��ml-1��Ũ������������һ���Ĺ�ϵ��С��4 ng��ml-1,��ʮ��ʮ�Ͽɷ������Ͳ�������������25��OH��D3���������嵥��ø�����£�øϵͳ����ϸ��ɫ��P450�����������ص��ף������Ȼ�����ת��Ϊ1��25��OH��2D3(1,25-dihydroxy vxtamin D),����ά����D������������ʽ���ֽ�����Ϊ���ء������÷�ʽ�������̴��༤�����ơ��ڰ���֯�ж��������壬1��25��OH��2D3�������γɸ������ڣ���ϸ���˻�Ⱦɫ�����ϣ�ͨ��DNAת¼���úϳ���ʹRNA��mRNA������ת��Ϊ�����ʣ�1��25��OH��2D3��Ѫ�����ɷ�����Ϊ52��00�ĵ���������������֯����С�����ǡ����ȣ�������Щ��֯�м���1��25��OH��2D3�����壬������ҪVD�ĸƽ�ϵ��ף�Calcium binding Protein,CaBP��,˵��1��25��OH��2D3��Ӱ�졣�����������������1��25��OH��2D3��CaBP�����߾������ڷ�ʮ���ȵ��صĦ�ϸ���ڣ���ά����D�ս�����£�������ֹ�ȵ��صķ�ʮ�ڣ�Ҳ����֤��1��25��OH��2D3���ڸ�ϸ����������ֻ��йء�
ͼ5-7 25-��OH��D3��1a��25��OH��2D3�Ľṹʽ
������1λ�Ȼ���ø��24λ�Ȼ���ø�����ƣ�ΪѪ��ˮƽ�����ơ�������Ѫ��Ũ���£�9.5mg%������1���Ȼ���ø��24λ�Ȼ���ø���л��������Լ��ܺϳ�1��25��OH��2D3Ҳ�ܺϳ�24��25��OH��2D3��Ѫ��Ƶ�ʱ���̼�1λ�Ȼ���ø���ƶ�ʱ���ƴ�ø���ɴ��Ե���1��25��OH��2D3�ϳ�֮����1��25��OH��2D3�ϳ����࣬24��25��OH��2D3�ϳ����٣���Ѫ���⣬������������Ӱ��1��25��OH��2D3���״�����أ�Parthormone,PTH���������أ�Calcitonin,CT������ʮ��ʮ���ض���ʹ�����ࡣ��Ϊ2���Ȼ�������Ҫ��֯����������������֤���ǡ�̥ʮ�̡��������ƾ��д˹��ܡ�
1��25��OH��2D3�ķֽ��л��24��25��OH��2D3��;�������ơ�24λ�Ȼ�����ɽ�һ��������24λ�����Ȼ��23λ�Ȼ������������ѡ�26-C��27-C������CO2ˮ��ʮ��ʮ��л����ά����D3-23����(Calcitroic acid)��Ҳ�ɲ���������������ά����D�ķֽ��л��Ҫ�����ڸ��ڣ��������л����ʮ�뵽��֭�У��ڷ�ά����D�ȴ�Ƥ���е��������ڷֽ⡣25��OH��2D3��1��25��OH��2D3Ҳ��������������ʽͨ�������γɸγ�ѭ����Ӵ������ʮ�����ڷ���������48h��30%�ļ����Ӵ������ʮ������2-��4%��������ʮ����
5.3.3����������
��1��ά��Ѫ�����Ũ�ȵ��ȶ� Ѫ��Ũ�ȵ�ʱ���յ���״�����ط�ʮ�ڣ������ͷ���������ϸ����������PTH���̼�1λ�Ȼ�ø������24λ�Ȼ���ø�⣬����ʹ�״�������ʮ����������С���������ա��ڹ���PTH��1��25��OH��2D3Эͬ���ã����ƴӹ��ж�Ա��������С����1��25��OH��2D3�ٽ��Ƶ����ա���������;��ʹѪ�ƻָ�������ˮƽ���ַ�������PTH�ķ�ʮ�ڼ�1��25��OH��2D3�ĺϳɡ���Ѫ�Ƹ�ʱ�̼���״��Cϸ�������������أ���ֹ�ƴӹ��ж�Ա����������ʹ�Ƽ��״�������ʮ����С��������Ϊ�������գ���Ҫ�������ơ������ǡ�1��25��OH��2D3��Ѫ����ʱ��8mg%���£����̼�1��25��OH��2D3�ĺϳɣ��ٽ�С���Ըơ������ա�����PTH���μӷ�Ӧ�����Ըƴ�������ʮ�����ײ���ʮ�����Ӷ�ʹѪ���������������������϶࣬ʹѪ�ָ�����ֵ��
��2���ٽ����м���ʮ��ʮ�����Ƶ����� 1λ�Ȼ���ø����Ѫ���и���Ũ�ȼ���ʳ�и���������Ӱ���⣬���ܼ� �ص�Ӱ�죬ͣʮ����ĸ�Ů1��25��OH��2D3Ũ�ȼ��ͣ����й���������֢״��
�ڻ����ڼ�1��25��OH��2D3Ѫ��Ũ����������ʮ��ʮ�ڼ�����������ʮ��ʮ��ĸ���ָ�������ˮƽ��24��25��OH��2D3֮ˮƽ��֮�෴���������½�����ʮ��ʮ��ָ���������̥ʮ��Ҳ��1λ�Ȼ���ø���ڻ����ڼ���������Ҳ�ܺϳ�1��25��OH��2D3��ʮ��ʮ��Ҳ��1��25��OH��2D3�İ���֯����ʮ��ʮ�иƵ�ˮƽֱ�ӹ�ϵ�����м���ʮ��ʮ�ڼ�ĸ�ɴ������Ĺ��н��������ά��̥��Ӥ������������ά����D��Ӧ�����ߣ��ڶ�ʮ��ʮ���ֿ����»�øƣ�ά����Dȱ���ߣ����ָֻ������ϲ
��3��1��25��OH��2D3���û��� 1��25��OH��2D3��С������Ϊ�յ��ϳ�CaBP.1��25��OH��2D3��С��ϸ���������γɸ���ʮ�����ϸ����Ⱦɫ���ϣ���ʹCaBP����ʹRNA(mRNA)�ĺϳɣ���mRNA�ڰ�����ת¼ΪCaBP�����ֵ��״�ʹ������ͨ����ʮëʮˢ״Ե��microvillus brush border���������ڳ�ϸ�����������������λ��ͨ��Na+��Ca2+��������-���Ĥ��(basal-lataaral mebrace)��1��25��OH��2D3����С��Ca2+����������������С������һ���ġ�1��25��OH��2D3Ҳ�����ڵ�Ѫ���Ƽ���ʳ�и�ȱ��ʱ�����ƴӹ��ж�Ա���������ڹ���δ������CaBP,1��24,25��OH��2D3���Դٽ�С�����ոƣ������ܴӹ��н��ƶ�Ա����������1��25��OH��2D3�Թǵ����û������С�����Dz�ͬ�ģ���Ŀǰ���������
�ǵĿ��ﻯ���õĻ�����δ����������1��25��OH��2D3��ȱ��ά����D�Ķ��P��ʮ�壬�����������ڹ��п����ʵij���������������Ȼ���������ά����D��л���ﵫ������δ�ҳ��ԹǵĿ��ﻯ�����������ߡ����ֽ���ֻ�˽ά����D�ٽ��������գ��ֿɽ����״ӹ��ж�Ա������ʹѪ���ơ��״ﵽ����ֵ����ʹ�ǵĿ��ﻯ�������ϸ��¡�
5.4.3����Ҫ��
��1��ά����D��Ӫ��ָ��Ѫ���м�ʮ��ʮ����ø���ӱ�ά����Dȱ������ʮ��ʮ֢״����Ҫ��һЩ���������ӷ���Ҳ��ά����Dȱ���ij����йأ��������Ƿ�����ʮ��ʮ�ģ���5-7����Ѫ����Ca��P�ij˻���mg%��,����ֵΪ36��40�����Ͳ��������������ߵij˻���30���¡�
��5-7 �����˼�����Ѫ���ʮ��ʮ����ø���ơ���Ũ��
| Ѫ���ʮ��ʮ����ø(���ϵ�λ��ml-1 (���ʵ�λumol��mm-1��L-1) | Ѫ��Ca(mg%) | Ѫ��P��mg%�� | ||
| ����Ӥ�� | 5��15 | 26��80 | 10 | 5 ~ 8 |
| ������ʮ�� | 3��5 | 16��26 | 10 | 3��4.5 |
| ���Ͳ��� | >20 | >115 | 8��9 | 3 |
| ������������ | 15 | 80 | 9 | 2��3 |
| ��״�����ع��� | 4��20 | 20��120 | 12��16 | 2��8 |
| �������� | 2 | 10 | 10��12 | 4��5 |
| ����ʮ��ʮ���ף�Paget's�� | 50 | 268 | 10 | 4 |
| �ɹ�ϸ���� | 30 | 160 | 10 | 4 |
����ע��1mgά����D��,�ⶨ��Ѫ����ֵ,��5����ٲⶨ����ʱѪ����ֵ,����ֵ����,���Ϊά����Dȱ����ָ�ꡣ�ڷ���������24h��Ѫ��Caֵ���͡�����������Ҳ��һ����������Ͳ��ķ�����
������ͬλ�ص���Ϸ����ⶨ25-(OH)D3����1��25��OH��D3��������Ѫ��25-(OH)D3Ũ��Ϊ15.2��5.6ng��ml-1����С��4ng��ml-1����ʮ��ʮ���������Ͳ�������������1��25��OH��D3Ѫ��������ˮƽΪ3��6ng��ml-1���������ⷽ�湤�����࣬������������������ˮƽ��δȷ����
��2��ά����D��Ҫ�������չ�����Ƥ���ɲ���ά����D�������Ӧ�貹�������չ������Ӱ�죬������Աδ�õ��չ�����1��3����֮�ã�ÿ�����ά����D��25��OH��D310ug����ά��Ѫ��25��OH��D3������ˮƽ���ӳ������ഺ�ڹ�Ӧ��Ϊ10ug��400IU���и�ʮ��ʮĸ10ug�����������������У������ж�ʮ̬ƽ�⣬�������Ͻ����ؽ�����ʮ��Ҳ��Ҫһ������ά����DԼΪ5ug��
5.3.5����Դ
5.3.6����ʮ��ʮӦ�ü�������Ķ�ʮ��ʮ
ά����D������ά����D3ȱ������1��25��OH��2D3�ɷ������в�֢������ʮ��ʮ�Dz��������ܲ�ȫȱ��1λ�ǻ���ø�����ڲ��ܺϳ�1��25��OH��2D3�����������ȡ���������ƿ�ά����D3���Ͳ��������Ŵ����أ��״�����ʮ�����ࣻ�ۼ�״������ȱ��֢�����߲����ڵ�Ѫ��Caʱ����1��25��OH��2D3���ܿ�ά����D�����Ͳ���ά����D��Ӧ�������������Ͳ������ڴ�л�ϵ�ȱ�ݣ�����1λ�Ȼ���������ﲡ��ʹ�ñ��ͱ����ܵ��¹Dz���Ҳ����25��OH��2D3����������Ϊ1��g/�졣�˼���Ҳ����Ϊ���Ƽ�����
ά����D�ж������������������࣬Ӥ������50��g��200IU�������һЩ���Ե���Ѫ�ƹ��࣬�����ܲ�ȫ����ʮ���ж������������ϴ����˿ڷ�2000IU�ж����ڷ�5000IU�����ж����ڷ������ܳ���800IU����ά����D����ʱ��Ҫ���Ѫ��ˮƽ����Ѫ�����������ж�������ж���Ż�£�ʳ��������������߿���������ά����D��ʮ��ʮ������Ѫ����25��OH��2D3ˮƽ�ߴ���1��25��OH��2D3�뵰�������ϣ����1��25��OH��2D3���ܽ���ϸ����Ҳ��������ƸƵ����ռ���Ա�ǸƵ����ã����Ѫ��ˮƽ�ߣ���ʹ�������༰�������ƻ�������ά����D����ʱ���õ��ż���Ա�ǸƵ����ã����Ѫ��ˮƽ�ߣ���ʹ�������༰�������ƻ�������ά����D����ʱ���õ��ż���Ƥ�ʼ����Լ���Ѫ��Ƶ�ˮƽ��
�ж�ʱ������ʮ��Ca�������Ѫ�ƹ��߷������磬��ƹ������γ�����ʯ��
ά����D��25��OH��D3���Դ��棬ά����D����ʱ��һ��Ϊ1��4���£��еĿɴ�18����֮�á�ά����D��л��Ҳ�ɲ����ж����������������˥�ڶ̣��ж�ʱ��Ҳ�϶̣�25��OH��D3�ɴ����ܣ�1��25��OH��2OH3�������ա�
5.4������E
5.4.1��ά����E�Ľṹ��ʮ��ʮ��
ά����E��VitaminE��VE���������ӣ�Tocpherol,T������ϩ�����ӣ�Tocotrienol,T3�����ܳơ���Ȼ�繲��8�ֻ��������һ��ɫ��������ֲ���IJ�����G-3��T����������ǰ�߲���3�࣬7�༰11λ��˫��������ɫ�������ϵļ�λ�ü���Ŀ֮��ͬ���в�ͬ���ͣ�������ʮ��ʮҲ��ͬ���Զ���������ʮ��ʮ�Ԧ�-TΪ��ߣ���5-8��5-9����������������Ό���������Ԧ���Ϊ��ߣ�������С����ά����E���������������ԳƵ�C�������γɹ�ѧ�칹�壬����ʽ�������ʮ��ʮ��Ϊd����֮�롣ɫ�����ϵ�OH������NH2���棬����Ӧ�͵�ά����E����ͬ�������ʮ��ʮ����CH-NH2����OH����»���͵�������������ʮ��ʮ�������ͬ��ɫ���������������������ʮ��ʮ������ģ��������٣���ʮ��ʮ�ͣ�����λ�ò�����Ҫ�ġ�
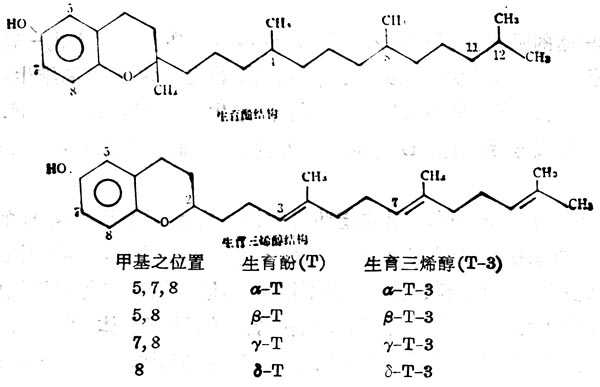
ͼ5-8 8����Ȼ���ά����E�Ĺ���
��5-8 ��ͬ���͵�ά����E����������������ʮ��ʮ���Ԧ�-ά����EΪ100%��
| �����ʮ��ʮ | |||
| ������̥���� | ������Ѫ | ����Ӫ���� | |
| ������E | 100 | 100 | 100 |
| ��-����E | 25��40 | 15��27 | 12 |
| ��-����E | 1��11 | 3��20 | 5 |
| ��-����E | 1 | 0.3 ��2 | - |
| ��-T-3 | 29 | 17��25 | - |
| ��-T-3 | 5 | 1��5 | - |
��5-9 ���֦�-ά����E�������������ʮ��ʮ
| ��ά����E������ | �����ʮ��ʮ��ÿmg�൱��IU���� |
| d-��-����E������ | 1.00 |
| D1-��-����E | 1.10 |
| d-��-����E������ | 1.36 |
| d-��-����E | 1.49 |
| D1-��-����E�������� | 0.89 |
| d-������E�������� | 1.21 |
ά����E����Ϊ����������Ϊ���ȼ�Fe3+,Cu2+���ٽ�������ʮ��ʮ��Һ�л���������½��ȶ�����ʽ������ʽ�ȶ������۲�Ʒ��Ϊά����E��������ӹ���ʳ����ʮ��ʮ�ƣ����Ư�����ж����ƻ���ʳ�ᆳ����Ҳ����ʧ�����ڵ�ʮ��ʮ�Ȼ�����½��пɼ�����ʧ��
5.4.2����л
ά����E�������������ʽ�ռ��������20��40%��������������һ����ˮ��Ϊ����ʽ��һ������Ϊ��ʽ�����Ա���ʱ����mg�ƣ������ʼ��͡������͵�ά����E�������������²�𣬵�ϸ���ɽ������֡������-ά����E���-ά����E�����������ƣ�����֯������-ά����E�����ޡ����Ԧ�-ά����E�������ʮ��ʮԼΪ��-ά����E��10%����Ҳ����Ϊ��35%��
ά����E��������֬��һ����Ӱ��֬����������Ӱ��ά����E�����ա������ɦ�֬��������ġ�Ѫ����֬������ά����EŨ������Ӧ�Ĺ�ϵ����֯��ά����E����������ȡ���Ķ������仯�����������ά���ز�ͬ������ά�����ڸ�����֯�У������⣩����һ������ֵ����֯�ϵ�ά����EΪ����ʽ��������ʮ��ʮ�١����´��塢ʮغʮ�輰ѪС����Ũ�����ϩ֬�������������ά����EҲ�϶࣬Ѫ��Ũ����֬�������仯����ѪС��Ũ�ȳ��ܼ������ı�����֬������ϵ�����ԣ���ѪС��Ũ����֬�������仯����ѪС��Ũ����������ı�����֬������ϵ�����ԣ���ѪС��Ũ����ΪӪ��ָ���Ѫ���ж�ȷ��֬����֯���μ�����Ϊά����E���Ĵ��泡������ϸ���ڵķֲ����ڸ�������������Ĥ��࣬�������Լ�����״ĤΪ��࣬��ϸ���ж���Ĥ�ϣ���Ϊ���ͣ���5-10����������ȡ��ͬ���͵�ά����E��Ѫ����ά����E��ӳ��ȡ������������������������ߡ���ĸʮ��ʮ��Ӥ��Ѫ�����Ц����ö������ͣ���ʽ��ʮ��ʮ���Ц����á����������͡�ιά����Eȱ������ʳ��Ѫ�������е�ά����E���սߣ������Ϊ�����������ļ��ڵģ�֬����֯�е��������������ڿ����ж��ֲ�ͬ�Ĵ�л�⣬һ�����ױ����ã�һ�ֲ�������ʧ��ϸ��Ĥ���߿������ں��ߡ�
��5-10 ��ʮ����֯�Ц�-ά����E����
| ������ | ��ʮ��ʮ��ά��ʮ��ʮ���� | ||
| ��g��g��֯-1 | mg��g֬��-1 | ��g��g��֯-1 | |
| Ѫ�� | 9.5 | 1.4 | |
| ��ϸ�� | 2.3 | 0.5 | |
| ѪС�� | 30.0 | 1.3 | |
| ֬����֯ | 150.0 | 0.2 | 2.4 |
| �� | 7.0 | 0.3 | 0.5 |
| �� | 13.0 | 0.3 | 3.5 |
| ���� | 19.0 | 0.4 | 2.6 |
| ʮ��ʮ�� | 11.0 | 0.6 | |
| ��ʮ�� | 9.0 | 0.7 | |
| �� | 20.0 | 0.7 | |
| ��ʮ��ʮ�� | 132.0 | 0.7 | |
| ʮغʮ�� | 40.0 | 1.0 | |
| ���´��� | 40.0 | 1.2 | |
ά����E��������ά����E������������Ԥ�������ƴ���ά����Eȱ�����µ���ʮֳ������Ӫ��ʮ��ʮ����ήʮ����ά����E����û������Ч�ã��������������ϴ�������ʮ����
5.4.3����������
��1��ά����Eȱ����֢״���ݴ��������ά����Eȱ������ʮغʮ�費������ʮ��ʮ�ӣ�����ʮ��ʮ����ֲ����ʮ���ڣ�̥�������ա�ά����Eȱ����������ײ��䣬���м���������ԣ���ʳ����dz���ʮ�У��ļ�����ԣ�ͻȻ�����������������ά����Eˮƽ�ͣ���ϸ����Ѫ������ʮ�У�ά����Eȱ�������ϸ�����ʱ��̣�����ȱ���ĺ�����ƶѪ����
ά����Eȱ����������ࡢ���⼰ʮغʮ�費������֯�IJ������䣬����øҲ�иı䣬������֯�еļ������ἤø�����⼰������ʮ��ʮ������ø���Ȳ�ת��ø�������б�ͪ�ἤø���ȱ�ת��ø�����Ͷ�Ѫ��������ø����ʮ��ʮ����ø���ӣ�����������Щø��������֯���뵽Ѫ�������£�����������֯�仯�ϸ����ԡ�
��2����������
�ٿ��������ã�ά����EΪϸ��Ĥ����ϸ����Ĥ���ϵ���Ҫ����������ϸ��Ĥ�ɵ�������֬����ɡ�������Ƕ����֬����֮�У����Ӽ�ʮ��ʮ�������⣬���ṹ�����⣬Ĥ�ı������н�Ͻ��ɵ����ܵ��ף������ֵ�����ø�йء�ά����E��Ĥ��ɫ����������������������ڣ��벻����֬����[������������ϩ�ᣨ20��4�ᣩ]���ã�����4�࣬8��λ������֬����ϵͳ�����ɵ���ʮ�ɻ���֮�����ã��Ӷ�����Ĥ�ϵĶ�ϩ֬��������ʮ�ɻ��Ĺ�����ά����Ĥ������ʮ��ʮ��
��ҪNADPH������ø���ɳ�������ڳ���᪻�ø��Superoxide dismutase,SOD�������£���Ĥ�ϼ��丽����H+����������H2O2��H2O2�ֲ���Ĥ�������ڡ��ڰ����ڹ����Ĺ�����ø��Glutathione Peroxidase,GSH-Px����H2O2�ֽ⡣��Ĥ��H2O2��O3�γ��Ȼ���ʮ�ɻ���ά����E�����������ֽ�����Ľ��仹ԭΪά����E��ά����Eȱ������ʮ�ɻ�����ʱ��ά����E�ýߺ�Ĥ�϶�ϩ֬���������ʮ�ɻ��Ĺ�����ά����E������ø����������������ø��������ø��H2O2Ũ�ȵ�ʱ��Ч����С�����������ڿ�����ϵͳ����ά����Eȱ���������ø���ܻ������ӡ�
�羵�۲죬���⼰�ļ�������Ĥ������Ϊά����Eȱ���ij��ڲ��䡣����ϸ��Ĥ�����������̶���ĤΪ�������м����Ĺ��졣ά����E�������弰������Ĥɥʧ�����Աȣ�����ά����E���Իָ����ӹ�����Ҳ֤��ά����E��ϸ��Ĥ�Ĺ�ϵ��ά����Eȱ���ߵĺ�ϸ��Ĥ����������Ѫ����һЩø����Ĥ���ƻ�������֯����Ѫ���С�
ά����E��һЩ���ܿ����������������������
�ڶ�֬���л��Ӱ�죺ά����Eȱ���������ڿ��������ܼ��٣����༰Ѫ����֬����������ü�ǿ�����Ը���Ϊ����֬���лҲ�иı䡣ά����Eȱ���������������TG���ڸ�������73%��Ѫ��������35%�����̴��ڸμ�Ѫ���о����ӣ���֬�����ޱ仯����ɵ�֬����Ҳ�иı䡣ά����Eȱ��������е�ϩ�����ӣ�����֬�����������Լ��٣�C20��4�����ӣ�����TG���ӵĶ࣬��Ѫ���ж����������ӣ���������١�ά����Eȱ�������������Ĥ��֬����̼���ӳ�������������ǿ�����粹��ά����E����48h��ָ�������
���ڶ�����֬����ά����E��ϵ��������Ҳ���о���ά����Eȱ�����ﲻ�ۼ��Ե��̴��������֬������������ӡ�����ά����E�ߣ���������Ӳ�������ʼ��㷺ʮ��ʮ����ά����Eȱ������������������Ϊ�͡�������ʮ�������У���Ѫ֬�������ޱ�������Ѫ֬�����ߣ�ÿ�ո���600IUά����E8��16�ܣ�Ѫ��֬���ޱ仯��
�۶�˥�ϵ�Ӱ�죺Ѫ����֯��֬���������ˮƽ����������ӣ�ά����Eȱ������Ҳ�����Ƶ������ԡ��ļ�������ά�к�֬�ʣ�Lipofusin����ͬ���䲹��ά����E��Ҫ�ࡣ��Щ���ߴ�ά����E�Ե���лӰ�����ά����E��˥�ϵ����á�����ι�Բ�ͬˮƽά����E83�ܡ���ά����E�������ӵ�70�ܣ�ʮغʮ��Ҳ�ȶ�������Ƴ��˻�ʱ�䡣N����9��ʱ��࣬22�������٣�66��ʱN��ʧ��������ȡ��������ά����E��N��ʮ�����ȵ���Ҫ�١�Ѫ���ܵ��״�9�ܵ�44�������ӡ���ά����E�߱ȵ��߶�9��16%���Ժ�Ѫ���ܵ������͡�����ĩ����ά����E��Ϊ����֮121%��ά����E���ܼ������������ʷֽ��л���ٶȡ���Щ���겡ѧ����Ϊ˥�ϵĹ�������ʮ�ɻ���֬�ࡢDNA���������Ļ��ۣ��������Ÿ��Դ����ά����E�Լ���˥�Ϲ��̡�
�ܶ�ǰʮ��ʮ��������Prostaglandin,PG����Ӱ�죺ά����E�Ĺ����翹ѪС��ۼ��������˻����ܶ���PG�йء������ⷽ�湤�����ڽ����У���δ������ά����Eȱ������ѪС��ϳ�PG���࣬���⼰ʮغʮ�����֯�кϳ��ּ��١�ÿ�ռ���400��1200IU��ά����EҲ�ɼ����˵Ľ�ԭ�������յ���ѪС��ۼ���ά����E��������ͬ��Ч�á�ά����E���ܶ�Ѫ˨�����кô�����ʮ��ʮ��������ά����EʹѪС��ά����EֵΪ����ֵ3��������ѪС��ۼ����á�PG����C20��4���֬������ø�����γ��ǻ�C20��4�ᣬ��ͨ��������ø���ö��ϳɡ�ά����E�����ƻ�����ø���ã���ˣ�ά����Eȱ����������϶��PG������Ѫ?f��ȣ�ʹѪС��ۼ����ü�ǿ����Ҳ������Ϊά����E����֬������ø���ã�ʹC20��4�����Ȼ���20��4����١�
����Ҳ�Ƕ�ά����EӪ��״����ʮ�е���֯��ά����Eȱ�����ﻷ����ø�ܵ����ƣ�PGE2��PGF2�ϳɼ��٣�����Ϊ���ⷢ�����估C20��4�������ԭ�ڷ�ά����E48h��PG�ϳ����������ἡ�ἤø�����ָ��������ڷ�5�պ�Żָ���
�ݶ��۾���Ӱ�죺����Ĥɫ����Ƥ��֯��ά����EӪ��״���ر���ʮ�С�ά����Eȱ������ɫ����Ƥϸ����ϩ֬������٣�֬�����������ۣ���ø����Ŀ���࣬����Ĥ�������٣���������ε�Զ�˵�Բ��С�ݻ���Vesiculation������ά����Eȱ������6���£��ټ��Դ���ά����E������Ĥϸ��Ĥ����������ɫ����Ƥϸ�����к�֬�ʻ��ۡ���������Eȱ������һ�����ң�����������ξ���ʧ����̬�ϵĸı�Ͳ�������ά����A��Eȱ����ʹ��������Ĥ��״ϸ����ʧ�����߾�ȱ��ʱ�Ƚ�ȱ��һ��ʱ��ʧ�����ء�
�����ά����E���Լ��ٸ�O2�Ի�����������۾�����ά����������������ѳ�����O2�����ܲ����۾������ά��֯�γɣ�RLF����ע��ά����E��Ԥ����
�Ի�����Ⱦ�Ŀ������ã���������ؿɲ�����ʮ�ɻ���ά����E�ɼ����䶾ʮ��ʮ�����п�����NO2��O3��ʹ�����ˡ�����NO2��¶��ȱE�����ά����E���½��⣬����ָ������أ������ʡ�֬�༰������ʮ�ɻ�øϵͳ���������ӡ�����ά����E�߷�ά����E����������ָ���ı䡣ȱE������ά���ؿ��������Ρ����붯������������ʮ�ɻ�øϵͳ��
ά����E��ʮ��ʮ�ǰ���CCl4�����������˵�֬�ʹ�����Ҳ��һ�����������ã��Լ�����Ǧ�ж���һ���ϵ��ⶾ���á�
ά����E�Ŀ��������ڶ���������δ�϶�����ά����E���ƻ����������ӣ�����ʮ��ʮ����·�Ӧ�졣��θ��ά����E����������������ɱ�ά����C����Ч��
5.4.4����Դ
ά����E�㷺������ֲ��ʳƷ�У�����ʮ��ʮʳƷ��d����Ϊ����ֲ���ͣ�����ͼ�Ҭ�����˳��⣩��ά����E�����϶࣬��������ȶ�ϩ֬���Ậ����ƽ�С�ά����EΪ��ϩ֬����Ŀ�����������ά����E�ڸ���ֲ���������Ͳ�ͬ�����ͽ��٣������ͽ�ռ8��10%������ʳƷ�Ħ�-ά����E���������и����͵�ά����E�ֱ������5-11����5-12�С�
Ӱ��ʳ����ά����E���������غܶ࣬ʳ��ɷֱ������ֲ��ܴ���ij����Ʒ�����֡���ţʮ��ʮ�ڲ�ͬ������Ҳ��ͬ��ά����E���ȶ����ڴ�����������ж�����ʧ��ը����Ƭ����ʮ��ʮ�д���2�ܣ���ʧ48%��ֲ�����ڴ����������ʧ���٣�ʮ��ʮ�Ƽ����ʱ��ʧ�϶ࡣ���Ư��Ҳ�����ƻ�ά����E��ʳ������������Ӵ���ά����E��ʧ�϶ࡣ
��5-11 ʳƷ�Ц�-ά����E270�ĺ�������g��g-1��
| Ӳ���� | ���� | 27065 | ˮ�� | ƻ�� | 33 | �߲� | ����� | 16 | |
| ������ | 210 | ���� | 2 | �ʶ� | ��1 | ||||
| ŷ��� | 72 | ���� | 3 | ��ʮ��ʮ���� | 10 | ||||
| ���� | 5 | ���� | 2 | ʮ��ʮ�ܲ� | 20 | ||||
| ɽ���� | 10 | �� | 13 | ���� | 4 | ||||
| ���Ӽ����� �� | ���� | 17 | �� | 5 | �㶹 | 3 | |||
| ���� | 7 | ��ݮ | 12 | ���� | ��1 | ||||
| �ף��֣� | 1 | ţ�� | 6 | �ܲ�ӧ | ��1 | ||||
| �ף��ף� | 10 | ���ࣺ | ���� | 5 | ���� | 22 | |||
| ���� | 11 | �� | 3 | ���� | 25 | ||||
| С�� | 117 | ����Ŀ�� | 9 | ���� | 24 | ||||
| С����ѿ | 5 | �㣺 | �� | 2 | ţʮ��ʮ������ | 12 | |||
| ȫ����� | 0.7 | Ϻ | 9 | ţʮ��ʮ��� | |||||
| ���� | 11 | ||||||||
��5-12 ֲ������ά����E�ĺ�������g��g-1��
| ��-T | ��-T | ��-T | ��-T | ��-T-3 | ��-T-3 | ��-T-3 | |
| Ҭ���� | 11 | ||||||
| ������ | 11 | ||||||
| ������ | 159 | 50 | 602 | ||||
| ����� | 100 | ||||||
| ������ | 189 | 214 | 21 | ||||
| �Ͳ��� | 236 | 380 | 12 | ||||
| �컨�� | 396 | 174 | |||||
| �ƶ��� | 79 | 593 | 264 | ||||
| �������� | 487 | 51 | 8 | ||||
| ������ | 1194 | 172 | 260 | 271 | 26 | 181 | |
| ����� | 211 | 316 | 143 | 32 | 286 | ||
| ������ͣ����� | 139 | 252 | 63 | ||||
| ������ͣ�Ӳ�� | 108 | 272 | 32 |
5.4.5����Ҫ��
��1��Ӫ������ָ��
�ٺ�ϸ����Ѫ���飺��ϸ����2��2.4%H2O2��ʮ��ʮ����Ѫ������Ѫ�쵰����������ˮ��ʮ��ʮ���ܳ�����Ƚϣ��ðٷ�������ʾ����ֵ��Ѫ��ά����Eˮƽ��һ���Ĺ�ϵ��
��Ѫ������ϸ����ά����E�ⶨ����ϸ����ά����E������ƽ��ֵΪ230��13��g%,Ѫ��Ϊ984��914��g%����ϸ����ά����E�仯��С��Ѫ����ά����Eˮƽ����0.5mg%Ϊȱ������Ѫ��ֵ����֬����ء�Ѫ֬��ʱ��Ѫ��ά����E�ͣ�ά����E���ܲ���ȱ����������Ѫ��ά����E��֬���������ʾά����E��Ӫ���������ÿ��֬��ά����E�ĺ�����������0.8mg�����õ�ά����EӪ������ָ��������5-13��
��2��Ӱ����Ҫ��������
��ά����C��ά����E�Ĺ�ϵ��ά����C��ά����E���п��������ã���ά����EΪ֬��ʮ��ʮ�ߣ���ֹ����Ĥ��֬�����������Ч��������Эͬ���á�ȱE��������ά����C��ʹѪ��ά����Eˮƽ���ߣ������ܼ���֬����������ϸ����Ѫ���ü�GSHˮƽ��ά����C��ʮ��ʮȱ��������֯��ά����Eˮƽ����50%���������˵��ά����C�ɽ�Լά����E���������ά����C������֮�෴�����Լ���ά����E������������ι��0.41IUά����E������ÿ�ղ���2��10mgά����C����ά����C���ϸ����Ѫ����֬����������ü�ǿ��Ѫ��ά����E����ϸ��GSH���١�ι0.8IU��ϸ����Ѫ��֬�����������ò���Ӱ�죬˵�������ά����C�ܼ������ڿ���������������Ӧ�����ά����E��Ҫ����
��5-13 ά����EӪ������ָ��
| Ѫ��ά����Eˮƽmg% | ��ϸ��H2O2��Ѫ% | |
| ȱ�� | ��0.50 | ��20 |
| �� | 0.50��0,70 | 10��20 |
| �ɽ��ܵ�ˮƽ | ��0.70 | <10 |
����ʳ��PUFA�ĺ�������ʳ��ά����E��PUFA��ֵӦΪ0.4��0.5��
��������������Խ�Լά����E��
��ҩ���Ӱ�죺Ůʮ��ʮ����ҩ����˹���ֶ�����E��Ҫ����
��3����Ӧ�� ���������Ӧ�����ڱ�5-14����Щ����Ϊÿ�չ�Ӧ15IUΪ��ˮƽ������ÿ�չ�Ӧ30��45IU��
��5-14 �������о�ίԱ�ᣨNRG�������Ӧ��
| ���� | ������أ�kg�� | ά����E��Ӧ����IU��d-1�� |
| Ӥ����0��0.5�� | 6 | 4 |
| 0.5��1�� | 9 | 5 |
| ��ͯ��1��3�� | 13 | 7 |
| 4��6�� | 20 | 9 |
| 7��10�� | 30 | 10 |
| �� ���ഺ�ڼ���ʮ�� | 70 | 15 |
| Ů���ഺ�ڼ���ʮ�� | 58 | 12 |
| �и���ʮ��ʮĸ | 15 |
5.4.6����ʮ��ʮӦ�ü�������Ķ�ʮ��ʮ
��1����ʮ��ʮӦ��
��֬�����ղ�ʮ���Ļ���ҲӰ��ά����E���գ��ڷ���Ч��Ӧע��ά����E��
��������������ѣ������������ƣ�Ӧע��ά����E������Ϊÿ������15mgά����E��������
����Ъʮ��ʮ���л��ߣ�ÿ�ո�300IUά����E�����Ӽ�����ά����E����������֫ʮ��Ѫ����
�ݼ�����ʮ�ɻ��Ի�������������Ÿ��Դ����ά����E�Լ���˥�ϵĹ��̡�ά����E�ɼ����Ԥ��O2��O3��NO2��CCL4����ʮ��ʮ�Ի������
��2��������Ķ�ʮ��ʮ �ڶ��������У������ά����E�������������ż�״�ٹ��ܣ���֬�����ӡ�ά����EҲ�ɸ���ѪҺ���̣������������ά����Eȱ�����ά����E��л�����E������ά����K�ṹ�����ƣ��������ά����K����Ҫ�������������������ϣ���������������Ѫ��λ�����������������צ�������Ѳ�������ά����K1��3�ռ�����ֹ����Ѫøԭʱ��Ҳ�ӳ���
��ʮ��ʹ�ô����ά����E��δ�������ж�ʮ֢״����10��000��ÿ����ȡ200IU�����ܣ�5000��ÿ�տڷ�1000IU��11��֮�þ����ж�������61������ȸ����������������ʡ�Ƥ��ƣ�͡�����28���շ�100��800IU3��֮�ã�ѪҺ��飬ѪҺ���̣������������״�ٹ��ܾ���������ʮ��Ӱ�졣
5.5������K
5.5.1���ṹ��ʮ��ʮ��
ά����K��VK����������ʽ����ֲ������ΪҶ������Phylloquinone,VK1�����ڶ����з����ά����K2(Menaquinone,VK2)��������ϵ������ϩ������Ŀ���ȣ�����ϸ������������һ���ͣ��ֿɼ��ΪMKn��n���������ϩ������Ŀ��M4��5��7��ά����K1�����ʮ��ʮ��ȡ�MK1ֻ��ά����K1��1%��ʮ��ʮ��MK10Ҳֻ��30��49%�Ļ�ʮ��ʮ��ά����K3��Menadione������ʽ�ϳɲ���ڲ�ʮ��ʮ�༰�������ڿɱ��MK4��ά����K1,ά����K2��ά����K3�Ľṹ��ͼ5-9��ά����K1�����˹��ϳɣ�����Ȼ������Ϊl�ͣ��˹��ϳ��߿�Ϊ�����ߣ����ߵĻ�ʮ��ʮ��һ���ģ���ά����K3��3λ����������Ҳ�����ʮ��ʮ��ά����K����������ά����K3������������ά���ػ����������Σ���Ϊˮ��ʮ��ʮ�ģ�����Ϊ�����õ��Ƽ��������������͵�ά����K����Ϊ������ƻ�����Щ����������������������нϸߵ�ά����K��ʮ��ʮ�����Թⲻ��ʮ�С�ά����K3��ά����K1��2��3λ����������䲻����ˮ�����Թⲻ����ʮ�У������ڿɱ�Ϊ��Ӧ��ά����K��
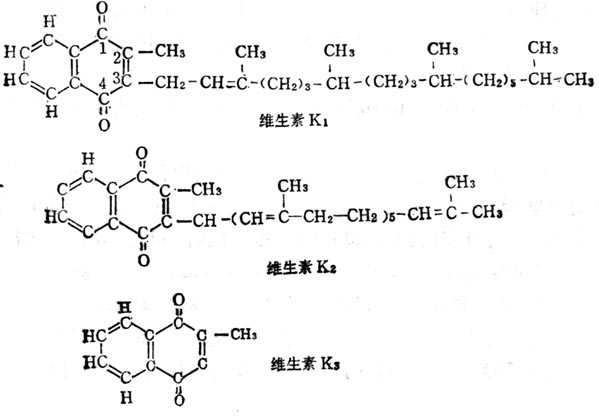
ͼ5-9 ά����K1��K2��K3�Ľṹʽ
5.5.2����л
��ʳ���ά����K�д�ʳ�������ģ�Ҳ�дӳ���ϸ���ϳɵġ�ά����K1��ά����K2������������֬��ʮ��ʮά����һ������Ҫ��֭����Һ������ʮ��ʮ�������ϣ����ܰ�ϵͳ���䡣������Լ��������10��70%���˻���ڷ�������ҩ��������ά����K1��20min��Ѫ�����ѳ���ά����K1,2h�ﵽ��ʮ�塣��48��72h��Ѫ��Ũ�Ȱ�ָ���½���1��5ng��ml-1�������ʱ�䣬����ʮ��ʮ����ת������֬�����У����������ڣ���VLDL���ϣ���ͨ��LDL������֯����ΪVK����Ҫ����֯��ע��ά����K1h��50%�����ڸ��ڣ��ڷ�VK2h��20%�����ڸ��ڣ�24h�������ֵ�����������ࡢƤ��������֮����24h�����ӵ����ֵ�����½����������ά����K����ԼΪ8��44ng��g-1����ε�Ũ�ȵ���4.5 ng��g-1,��Ѫøԭʱ���ӳ���ά����K�����Ƚ�С��Լ50��100��g��ת���ʿ죬�����ÿ2.5h��ת��һ�Ρ����Ĵ�л��Ϊά����K������������л���γɦ�-���������������������ϣ�����ʮ��ά����K�IJ������Խ��Ц»�������γ�6`-�Ȼ��ἰ���-�������һ���ֽ�Ϊ4`-�Ȼ��ᣬ���������Ļ�����л���Щ��л���������������ϣ������ڳ���ѭ���У����������ʮ����
ά����K3�ڶ����������ת��ΪMK4�����������٣���Ϊ��ȡ����0.05~1��0%��ά����K3��Ҫ��л����Ϊ˫��ά����K3�����������������
5.5.3����������
��1��ά����KΪ�Ȱ����-�Ȼ���øϵͳ�еı������ء���-�Ȼ��Ȱ��ᣨ��-Carboxyglutamic Acid,Gla���ĺϳ���ϸ�������ڽ��У���Ҫ���йȰ����������Ϊ���ʣ�����Ҫ����������̼��ά����K������ά����KH2���������������ά���صı仯����ά����K-ά����K2��3���������ά����K-2��3epoxide,VKO��ѭ������ʾ����-�Ȼ������õĵ������˹��ϳɵ�����������Ȼ��Դʮ��ʮ���ף�����Ѫøԭ�����˹��ϳ����Ա���-��-��-��-����Ӧ����ǿ�������籽��-��-��-�Ӽ�����-��-��-��-����Ҳ�����á��˹��ϳ�������Դ����֮���о�����������������ʱ����Դ���Ħ�-�Ȼ����Ŀ�ʼ�ٶȼ��٣���Դ���״���ʱ�Ƴ���������-�Ȼ�����ʱ�䡣
��-�Ȼ��Ȱ��ᣨ��-CacbocyglutamicAcid,Gala���ĵ����ʻ��������γɺ���Gla���ڵ��Ȼ�������Ƽ���֬��ϵ���ʮ��ʮ��
Gla�����ʿ����������ɳ��������������֯�з������á�Gla���ֽ�����ղ���Ϊ����ʽ��Gla����Gla����������������ʮ����������������ʮ����Ϊ44��11��mol��g-1��������ͯ��ʮ���϶࣬5��ʱԼΪ100��mol��g-1�������Ժ���ʮ�������½�����15��ʱ������ʮ��ˮƽ��ʹ�ÿ������ߣ�����Glaˮƽ��Ϊ24��mol��g-1��������Ѫøԭʱ�������������2��3����Gla������ʮ����25%�����ܿ�������Ӱ�죬�ɴ������йǸƵ��ף�osteocalcin����ת������Щ����Gla����ʮ��Ҳ�б仯����������������ɲ��ˣ�����Gla����ʮ��������������50%���൱�ڹǵ�ת����֮3����Ƥ������ӲƤ������������ʮ��Ҳ���ӡ�
��2��VK��ѪҺ���̵Ĺ�ϵѪҺ�����Ǵ���֯���˺�ѪС���ƻ��������һϵ�е�ø����ʽ��Ӧ��ѪҺ���̹�����һЩøԭ��proenzyme���ĺϳ���ά����K�йأ��༴�����ǵĺϳ�����Ҫ�Ȱ����-�Ȼ�������Щøԭ�����Ӣ����������⣬����������˵���C��S��M��Z���������·��ֵĵ��ף����ǵ�1��40����������˳������ѪøԭͬԴ������C����ѪҺ���̣����ٽ�Ѫ��ά�����ܽ⣬�������ĵ���C����ʹ���Ӣ�����������S���Լ�ǿ����C�Ļ���������10��Gla������M���Դٽ���Ѫøԭת��Ϊ��Ѫø������Z��13��Gla����Щ�����˽�û����٣���Ҫ��һ�����о���
��Ѫøԭ�ĺϳɣ����ڸ�ϸ����������Ĥ�ϵ��γ�������������Ȼ���ٽ���һЩ�Ȱ���Ħ�-�Ȼ������ǻ���������Ѫøԭ��NH2ĩ�˵�7��8��15��17��20��21��26��30��332λ�õĹȰ����-�Ȼ������Gla��33λ��ĹȰ��ת��ΪGla�����֦�-�Ȼ���������ѡ�����ɰ����������˳�����£��������ڵ���ǰ����Ĥ�ϵ�λ���빹�����¡�1���ӵ�������Ѫøԭ��10��12Ca2+���ϣ�δ��-�Ȼ�����ֻ����һ������Ca2+��ϡ�
��3��ά����K��ǻ����к�Gla���ף�Bone Gla Protein,BGP���Ĺ�ϵ �ǻ����м��ֺ�Gla�ĵ��ף���ҪΪBGP��Ca����߽����ǸƵ��ף��ڹ�ϸ���ںϳɣ���ʮ�ڵ�ѪҺ����֯��Ȼ�ǻ����У�ռ�����ܵ���1��2%��Ϊ�ǽ�ԭ����10��20%���ǸƵ��׳����ڹǿ��ﻯ֮ǰ�����ܶ����ӣ���Ҳ���ӡ�����2���ƽ�ϵ㣬������Ϊ0.8mmol��L-1����ʹ��뱥�ͣ�����������������þ���ȡ���Ҳ����֮��ϣ��������ӽ��������ǿ�����������ڵ��ڸ��ڹǻ����г�����������ʯ��hydroxy apatite���ĺ��������á�Ҳ�м���˵��BGP�ĺϳ�Ϊ1��25��OH��2D3�����ڡ�BGP���ܵ���1��25��OH��2D���ƹ����ã�ʹ�����û�������һЩ�ǵļ����У�Ѫ����BGPˮƽ��������˵�������ܴٽ��ǵ��ؽ����ƵĶ�Ա��
����������ĸ����ά����K��������̥������������Ѫ������һ����˵������̥�����������У���Ҫά����K�Ĺ���ϵͳ������Ѫ������ϵͳҪ��һЩ����ĸ�彫��������̥����һ���̶�ά����K������ʮ�У����ܸ�����̥ʮ���Ц�-�Ȼ������ĺϳɡ�
��4������ ��С��ϸ���к�Gla�ĵ����ʣ�Ϊ���ܵ���0.2��0.7%��������Ĥ��ϵĵ��ף�Ҳ������ϡ�������С��ϸ���ڵ���������֮�йء�������֯�����ʡ�̥ʮ�̡�ʮغʮ�衢�ȡ�Ƣ���Ρ�ʮ��ʮ�ٵȶ���Gla�����ʣ����ò�������Щ��֯�缡�⡢���༰�ܰ�ϸ������δ���֡�����Щ����������ʯ�������Dz���Ƽ���ʯ��ʯ������Gla�ĵ����ʡ�������������֬�����Ƽ���ά�߿���û�к�Gla�ĵ����ʣ�������Ӳ���ƻ��߿��к���Gla�ĵ����ʡ�
5.5.4����Դ
����ά����K����Դ�ж����棺һ����ӳ���ϸ���ϳɣ�ռ50��60%��VK�ڻس������գ�ϸ�������ڻس��ںϳɣ�����Ϊ��ʮ�������ã���Щ����������������������ϸ��������Ӱ��ά����K�����롣��һ�����ʳ��������ռ40��50%����Ҷ�߲˺����ߣ������ʮ��ʮ�����࣬ˮ�������ຬ���͡�ʳ����ά����K�ĺ������ڱ�5-15��
5.5.5����Ҫ��
��1��Ӫ������ָ�����Ѫøԭʱ�䣬������Ѫ�ӵ�һ�����Ĵ���Ѫøԭ��ø��Һ�У��۲�������ʱ�䣬ԼΪ25��40s��������ʱ�䣺����Ѫ����ʱ�估�γ������ʱ�䡣������Ѫ����ʱ��Լ10min��
��5-15 ʳ����ά����K�ĺ���������g��100g-1��
| ����ʳƷ | ���� | ���� | ���� | �߲� | ���� | ˮ������ | ���� |
| ţʮ��ʮ | 3 | �� | 5 | ����� | 57 | ƻ���� | 2 |
| ʮ��ʮ�� | 35 | С�� | 5 | ������ | 200 | �㽶 | 2 |
| ���� | 30 | ȫ�� | 17 | ���ײ� | 125 | ���� | 1 |
| ţ�� | 15 | ��� | 4 | ���� | 129 | �� | 8 |
| ţ��ĩ | 7 | ��� | 4 | �㶹 | 19 | ���Ѹ� | 6 |
| ���� | 11 | ���� | 20 | ���� | 89 | ���� | 38 |
| ���� | 15 | ������ | 0 | �ܲ�ӧ | 650 | �ɿڿ��� | 2 |
| Ѭ���� | 46 | �컨�� | 0 | ���� | 3 | �̲� | 712 |
| ţ�� | 92 | �̶� | 14 | �Ϲ� | 2 | ||
| ���� | 25 | ������ | 5 | ||||
| ���� | 7 | ˮ��棨watu cres�� | 57 |
��2����Ҫ��ԼΪ2��g��kg-1����-1������Ϊ10��g��kg-1���ػ�0.1��g��gʳ��-1����������Ҫ��Ϊ25��g��kg����-1��
5.5.6����ʮ��ʮӦ��
������Ӥ���⣬����ԭ��ʮ��ʮVKȱ����Ϊ�������������ڳ��������������ϸ������̥����̥ʮ�̲�������֬�࣬��Ѫøԭ���е͵��������ܺ�����������ֵ�������������ֵ��10%��������������ʮѪʮ��ʮ������ˮ��ʮ��ʮ��֬��ʮ��ʮ��ά����K�Ƽ���������Ч�ػָ���Ѫøԭ������ˮƽ���������ʮѪ����Щ������Ҳ��ά����Kȱ����������Ѫøԭʱ�䳤������
Ԥ��Ѫ˨ʹ��˫���㶹�ع���ʱ����ѪøԭҲ���٣��ɾ���ע��ҩ����������mg�Ƶ�ά����K��ʹ��Ѫøԭ���������ڿ�ʼ�ϳɣ�24h�ﵽ����ˮƽ����ˮ��ʮ��ʮά����K3��������ת��Ϊ���ά����K���ٶ�̫����Ч�����á��ھ���Ӫ���У���ע��ʮҺ�����˿����أ�����ά����K�ڳ����ںϳɣ����˵���Ҫά����K����Ѫ������4���ڽ���������ֵ��20%������עҺ��Ӧ����ά����K0.03��1.5��g��kg����-1����������ʱά����K���ռ��٣�Ҳ�ɲ���ά����Kȱ�����ڷ�������������ʱ�������뵨��ͬʱ��������Щ�β������ڹ��ܲ�ȫ��Ѫ���Ӻϳɼ��٣�����ά����K����Ѫ������Ч��
5.6������B1
5.6.1���ṹ��ʮ��ʮ��
ά����B1�ֳ����أ�Thiamine VB1��,����ʮ��ʮ������ʮ��ʮ����������Һ�зֽ���ऺ����������֣�ͼ5-10�������뽹�����������ؽ����ᣨThiamine pyrophosphate,TPP�������Ȼ���ø��cocarboxylase��,�����Ӧ��ҪATP�μӣ��ṹʽ��ͼ5-11��TPP�����Ǵ�л�Ц�ͪ��������������á�
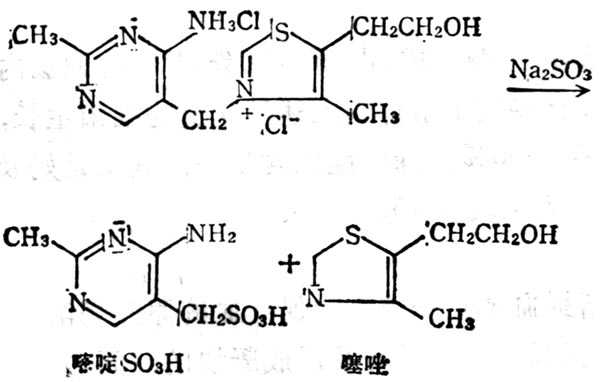
ͼ5-10 ά����B1��Na2SO3�ֽ�Ϊ��ऺ�����
ͼ5-11 �Ȼ���ø��TPP���Ľṹʽ
ͼ5-12 ��ɫ�ط���ʽ
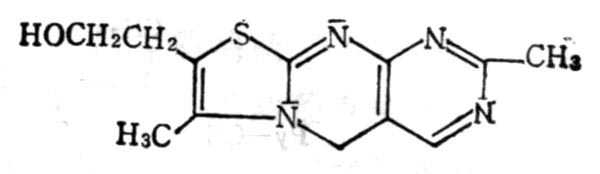
ά����B1������1g������1mlˮ�У����ھ�ʮ��ʮ���γ�1%��Һ�������������л���ý�С�PH7ʱˮ��Һ�����������ո�ʮ��Ϊ235��267nm���൱����़����������֡���PH1ʱ��ֻ��1����ʮ����247nm������260mn����һ���磬Ҳ�൱���������ɲ��֡��ڸ�ʮ��ʮʱ�������ڼ�ʮ��ʮ��Һ�У��dz������ƻ�������PH5�������ƻ����ڼ�ʮ��ʮ��Һ�У��������������������أ��ɽ�ά����B1����Ϊ��ɫ�أ�Thiochrome�����ݴ˿����ԲⶨVB1��ͼ5-12����
�˹��ϳɵ�VB1�������б������ض����Thiamin propyl disulfite,TPD����4����ȩ�������أ�thiamin tetrahydrofurfural disulfide TTFDD����������ˮ��������ø�ƻ����١��ڷ���Ѫ����֯���Լ���Һ��ά����B1��ˮƽ�Ϸ���ά����B1ʱҪ�������ɸ�10��������������Ҳ�����ա������Щ�����ﳣ����ʮ��ʮ��Ӧ�á���Щ��������������أ�pyrithiamino�������������п�ά����B1���ã�ǰ��ʹ������ά����B1���ڿսߣ����߿ɵ��³���ϵͳ������ϵͳ��ά����B1ȱ��֢״��
5.6.2����л
ά����B1��С�������գ�Ũ�ȸ�ʱΪ��ɢ����ʱΪ�������գ���Ҫ�����Ӽ�ATP��ȱ�������Ӽ�ATPø�����������ա���ˮƽʱ����Լ57.5%����ʱԼ26.2%��ά����B1���뵽С��ϸ�����ữ��������С��ϸ����ȥ��Ҳ��Ҫ����Ũ�ȵ������Ӽ�ATPø��Ҷ��ȱ����Ӱ��ά����B1�����ա�
����Լ��ά����B130mg��50%�ڼ����У��������ġ��Ρ��������н϶࣬����ά����B1��80%ΪTPP��10%Ϊ���������ᣨTTP��������һЩ���ص����ἰά����B1������������ø�����γ��������������ữ������ؽ������������ATP�����γ�TPP��TPP-ATP����ת��ø��TPP+ATP��TPP�������ؽ�����ø����TPP��TMP�ķ�Ӧ��
5.6.3����������
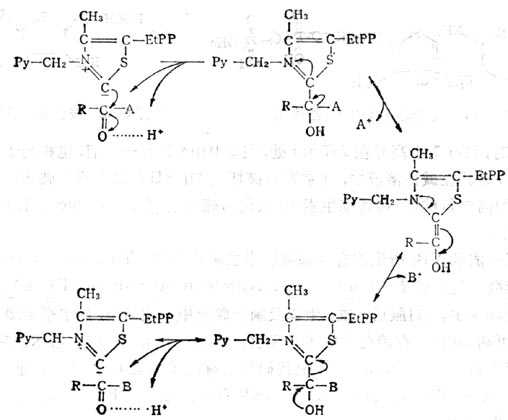
��1�����ؽ�����Ϊ�Ȼ�ø�ĸ�ø�������û�����ͼ5-13��
���������У���ͪ�����ȱ�Ϊ��ȩ���ų�CO2��
�������Ȼ���ĺϳɣ������Ȼ���Ϊ�ϳ�֧�������ᣨ�Ӱ��ᣬ��������ȣ����м��塣
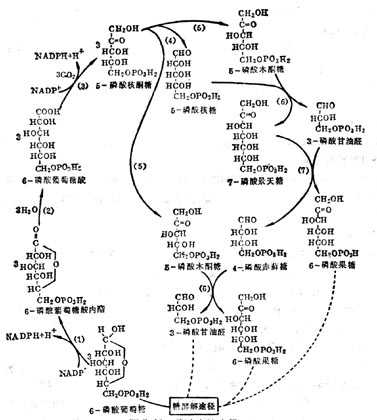
����Ϊ����������·��תͪø�ĸ�ø �ǵ������ͽ⼰���������Ƕ��������Ƿֽ��л����Ҫ;����������һ���־�����������·��л���ڸ�����ֻռ�ͽ⼰����;����8%�����ص���ͨ�����������ȷ�Ӧ����CO2���������ȩ��6-������ǣ�������Խ���ͽ�;���������Ϊ���������ữ��Ϊ6-���������ǣ�G-6-P������G-6-P����ø�Ĵ��£�������ˮ���ϳ�6-���������ᣬ�پ����������ȵȷ�Ӧ����5-�����ͪ�ᣬ��ת��Ϊ5-����ľͪ�Ǽ�5-������ǣ�ͨ��תͪ��ø����ҪTPP��Mg2+����ͪ���ϵĶ�̼��λת�Ƶ���һ��ȩ�ǵ�1-C�ϣ������Ӧ�ǿ���ģ������ѭ�����γ�7C��6C��5C��4C���ᵥ�ǡ�
ͼ5-13 ά����B1�������Ȼ�ø�����û���
ͼ5-14 ��������;��
�����л����Ҫ���������Dz������Ǽ�NADPH�������������Dz���5-������ǵ�Ψһ;�����������л�ж�����ø��6-��������������ø��6-��ø����������ø��������NADPΪ��ø����ԭΪNADPH����Ϊ�������෴Ӧ�Ĺ����壬��֬���ᡢ���̴�����̴��ĺϳɶ���Ҫ����������������Ӧ�Ƚ���ʢ��֬����֯����ʮ��ʮ��ʮ��ʮ�١���ʮ��ʮ��Ƥ�ʡ�ʮغʮ�輰����ȣ����ִ�лҲ�Ƚ���ʢ���Ŵ�ʮ��ʮ6-��������������øȱ���IJ��ˣ����ܽ�������������·ѭ����NADPHȱ����GSH�����£���ϸ�������ƻ���������Ѫʮ��ʮƶѪ��
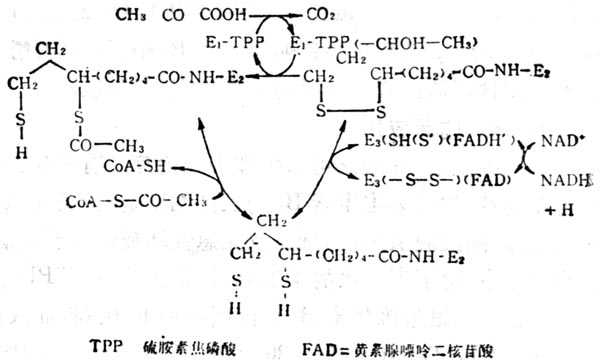
�ܦ�ͪ����ø�����ͪ�ᡢ��-ͪ����ᣩϵͳ�еĸ�ø������øϵͳ���������ã�ͼ5-15����
ͼ5-15 -ͪ����ø������
A �γɡ���ʮ��ʮȩ���м������Ϊ��-ͪ����ø1����-ͪ����ø�����ؽ�����Ϊ��ø�����ã����Ⱥ��γ��м����ø����ø���õ��м���Ϊ2-�����һ������ؽ����ᣬ��-ͪ�������м���Ϊ2-��1��3�Ȼ����������ؽ����ᡣ
B �������м���ת����ø2���������ϡ�
C ������ת������øA�ϣ���˫�������ᱻø3�����γ������ᡣ
��2�������������ϵ����� һ���嶯����ʹά����B1���ữ����ȥ���ᣬ��ʹ����Ĥ��ʮλ�ƣ�Na+������ʮ��ͨ��Ĥ��������������ʮ��ʮ��ά����B1�������ǽ����������ﻹ������������������в��ܿ϶����Դ��Թ������ã��������ڶ�5-��ɫ�������뼰��֬�ϳ���Ӱ�졣
��3������������ܵĹ�ϵ ά����B1ȱ�����������ʧ������ֱ�ӵ����ã���������ά����B1ȱ��ʹѪ���뵽��֮֯�����࣬ʹ����������Ӹ������أ�������ά����B1ȱ�����ļ�������л��ȫ��
5.6.4����ţ�⡢�Ρ����ȣ�ȫ���ס������߲ˣ�����ȸ���ά����B1����ʳ������Щ���ӿ�ʹά����B1����ı䣬�������͡����ֿ�ά����B1�����ӷֶ��֣�һ���ױ����ƻ���������ø��ǰ���ڵ�ˮ�㼰���������ڣ������صķֽ⡣���ߴ���ijЩ�����У���ά����B1�ķֽ⣬�����ȶ��Ŀ�ά����B1���Ӵ�����ֲ���У���������3��4˫�Ȼ������͵������йء���ά����B1��ȡ������Ⱥ��������Щ��B1���ӣ��ɵ���ά����B1ȱ����
�����ڳ�ȥ��Ƥ���ǵĹ����У�ά����B1��ʧ�ܶ࣬�����ʮ��ʮ�ӹ������۶�ǿ��ά�����������ʹ�����൱�ڴ���Ʒ�����Ӧ�����ҹ�ʳƷ��ҵ���ż������ߵ�ע�⣬����Ӽ��ʹά����B1��ʧ��
5.6.5����Ҫ��
����ά����B1Ӫ��״����ָ��һ����������ʮ����Ϊ��������ÿ�˼�����ʮ����ά����B1��ʾ֮����ϸ����תͪø��ά����B1Ϊ��ø��Ҳ���Բⶨ��ʮ���ά����B1Ӫ��״̬��
��ʮ��ά����B1��Ҫ��Ϊ1.26��1.47mg��1000Kj-1����������ȡ��������£�ÿ������ʮ����Ϊ40��90ug,����ȡ�����ӵ�2.1mg��1000Kj-1,ÿ������ʮ����Ϊ100ug�����ϡ�����ȡ�����͵�0.84 mg��1000Kj-1,����ʮ����Ϊ5��25ug�����������߿ɵ���0��15ug�����ϡ��ҹ������������Ӧ��Ϊ2.1mg��1000Kj-1�������������ص�Ч�ʽϵͣ�����Ϊ4.2 mg��1000Kj-1����ʮ��ʮ��ʮ��ÿ��1.2��1.5mg��Ů1.0��1.1mg,�и���ʮ��ʮĸ���乩Ӧ������0.3mg,��ͯ��Ӧ��0��6����0.3mg��6���¡�1����0.5mg,1��3����0.7mg,4��6��0.9mg��7��10��1.2mg����11��14��1.4mg,Ů11��14��1.2mg�����ƹ�����ά����B1��Ҫ�����ӣ���������˵��ά����B1ȱ���ߣ���ʮ��ʮ��ʮ�����������ӣ����������߿��Ե��²�ʮ����ʳʮϰʮ���߸��ų���ά����B1�����ա�
5.6.6����ʮ��ʮӦ��
����ά����B1ȱ�����������������ơ���Щ����ʮ��ʮ��л����Ҫά����B1�ļ�����Ҫ��ʮҩ������������Ҫά����B1�ľ�ϸ��ƶѪ��ʮ��ʮ�����б�ͪ������ø�����ͣ���֧��ͪ����֧��ͪ������ø�����ͣ����Ǽ�ʮ��ʮ����ʮ��ʮ�Լ��財������ϵͳ��ȱ��TTP��������Щ���������У��ò�����ˮ��ά����B1�������TPD��TTFD������ά����B1��Ч��Ҫ�á�
����ע�����ά����B1�����������ѹ�ֶ���������������ÿ�������أ�С��125mg������250mg������300mg����350mg��������Ƥ�¼��ڷ������������Ϊ1��6��40������ע��ÿ��������600mgʱ���ɲ����ж������˿ڷ������ά����B1��δ�������� ��ʮ֢��Ƥ�¡����⡢����������ÿ��ע����������1��200��ʱ����δ�����ж�������Щ�˽��ܴ������������ʮ��ʮ�ݿˡ�
5.7������B2
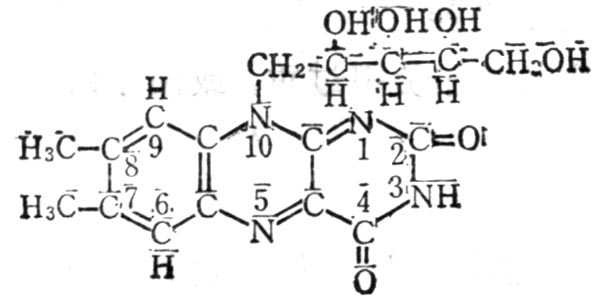
5.7.1���ṹ��ʮ��ʮ��
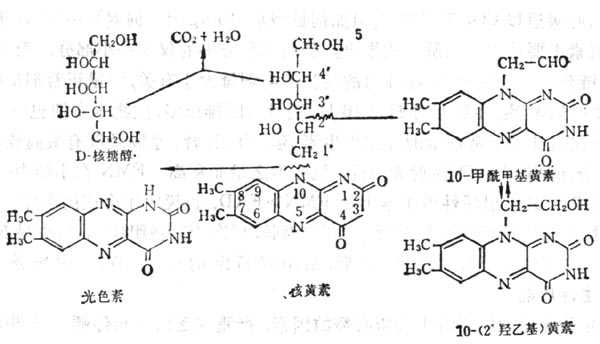
ά����B2�����˻��أ�Riboflavin,VB2�������Ǻ˴���7��8-��������ີ������ͼ5-1`6����Ϊ�ƺ�ɫ��״�ᾧ���ܽ�Ƚ�С����27.5��ʱ�ܽ��Ϊ12mg%���˻���5-��������ܽ⣬�����ڼ�ʮ��ʮ��ʮ��ʮҺ�������¾��ֽ⣬��ӫ��������λ�ǰ�������ֽ�����ɵù�ɫ�ء�����ؼ����������ء����������ڣ���������Ҳ�ɽ���ֽ�Ϊ���������صȣ�ͼ5-170��
ͼ5-16 ά����B2�ṹʽ
ֲ���ܺϳɺ˻��أ�����һ�㲻�ܺϳɣ�������ʳ�﹩�������ڲ�ʮ��ʮ���ﳦ���е�������Ժϳɲ�Ϊ�������գ���������������������Ҫ��
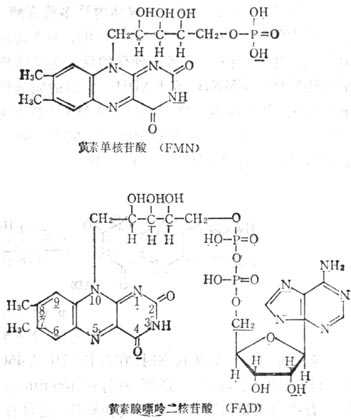
�˻��صĸ�ø�ж�����ʽ�������ص������ᣨFMN���ͻ��������ʶ������ᣨFAD����ͼ 5-18����
5.7.2����л
�˻��ػ��丨ø��ʳ�����뵰���ʽ���γɸ���������ص��ף���ʮ��ʮ�����е������������ڵ���ø��������øˮ��Ϊ�˻��ء���Щ���ص����ͷų�8��-�����ᣨ���װ�����鰱�ᣩ�˻��ؿ������գ���������Ч�á�
����С���������պ˻��ء�����������Ѹ�٣�������������ɱ�����ֱ����ȡ��Ϊ25mgʱ�����ȶ����˻���������ʳƷһ�����룬���������ա���θ���������Ҽ������������ɵ������ղ�ʮ����ά����B2�ڴ���Ҳ�����ա����յĻ����������С���죬�ͼ���ʱΪ�������գ���ҪNa+���ڼ���Ϊ0.2ugmolʱ�������������ʣ������ʱΪ��ɢ���ա�
ͼ5-17 �˻���5-����ķֽ����
ά����B2�����Ը�ø��ʽ������Ѫ����֯����ʮҺ�У���Ѫ������֯����Σ�ϸ���У�����ʽ�ĺ˻��أ�������ϸ��Ĥ��ϸ���ڡ��˻������������ʮ��ʮ�ǻ��ؼ��˻ƻ��أ�������ֹά��B2�ڻ��ؼ�ø��Flavokinase�������£�ת��ΪFMN�����ּ�ø��������ϸ�����ڣ����ڸμ�С���ڵĻ����ر�ߡ����ؼ�ø����ʱ��ҪATP������ʮ�ʮ���ӣ���Ҳ����ά����B2��������������������Ϊ���ż���ά����B2��������������������ض��Ǹ��ż���
FAD����FMNͨ��FAD�ϳ�ø�����γɵġ���ø�㷺��������֯�İ����ڣ�Ҳ��ҪATP����ʮ�ʮ���ӣ�FMN���������ﶼ����Ϊ�����˻��ظ������������FMN��FAD�������á�ά����B2ȱ������״�ٹ���ʱ��FAD�ϳ�ø�������ӡ�
ˮ��FMN��FAD��ø�㷺���ڸ���֯�С�FMN��ø�ڼ�ʮ��ʮ����ʮ��ʮ�ж������ã��ڸ�����ʮ��ʮ�����FMN�����ý�ǿ����ά����B2ȱ��ʱ������Ӱ��FMN����ø�Ļ�����FAD������øҲ�ܴ�ʹFADˮ�⡣���ڸβ����˵�Ѫ�������ߡ�
ά����B2����С���˹���������С���������ա���Ѫ��ά����B2Ũ�ȵ�ʱ������ʮ�������٣�Ϊһ��������Ѫ��ά����B2�ﵽ1��g��ml-1ʱ����ʮ�����ࡣά����B2��ȡ����С�����ڴ���ٷ������������55��g14C-һά����B2��10%��14C��������ʮ����3%�ڴ������ʮ����1%�Ӻ�������CO2��ʽ��ʮ��������������ӣ�15��g��ml-1���ϣ���10%14C�ӱ�����ʮ�������Ժ˻�����Ҫ������������ʽ��ʮ��������ϸ���ֽ�Ľ�ΪһС���֣����һ�״���Ƚ��ȶ����������ֽ⡣ά����B2�����ڵ�ת�����������С�йأ�������������ʱ��ÿ����ʮ��15��20��gά����B2�����������Ϊ16�գ�������ʱ��ʮ���࣬���������Ҳ�̡�
ͼ5-18 �˻������ʶ������ᣨFAD��
5.7.3����������
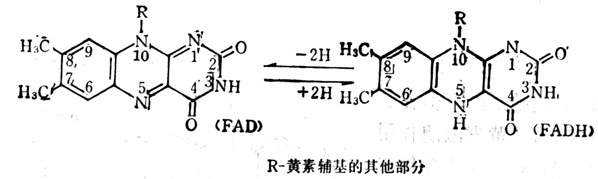
ά����B2��ά�ֶ���������������������أ�����ȱ��ʱ����ͣ�١���Щ�ֲ������ڿڽǴ���ά����B2��ȱ�����������á��ڻ���ʱ����ά����B2ȱ�������»�̥������������δ���������������������֤����Щ��������ά����B2ȱ���йأ�����FADԤ����
ά����B2��Ҫ�����ǹ��������������ø�еĸ�ø��FMN��FAD������ЩøΪ���Ӵ���ϵͳ�е�����ø������ø��ͼ5-19����ǰ�߽���ԭ�͵ĸ�ø�������ֱ�������ã����ɹ������⣬���ýϿ졣���������ķ�Ӧ�Ǵӵ����Ͻ�һ˫��ԭ��ֱ�ӵݸ�FMN��FAD���γɻ�ԭ�͵�FMNH2��FADH2�����ǵ���������������ϵ��������أ�����ֱ�ӱ������������������û������ڻ������ں�������FMNH��FADH����������NADPHҪǿһ����������ֱ���������ã���NAPHֻ�ܼ���������á�FMN��FAD�����ǽ���NADPH�ݸ�����ʹ����FMNH��FADH��
ͼ5-19 ����ø������
���ص��״�������״̬ʱ���ڲ���370��450nm�����нϿ������շ壻���ڻ�ԭ̬ʱ����450nm�ķ���ʧ����Щ���ص��׳����ƺ������⣬�����н������ӣ��������⡢п����Ҳ����Щø����Ҫ�ġ�
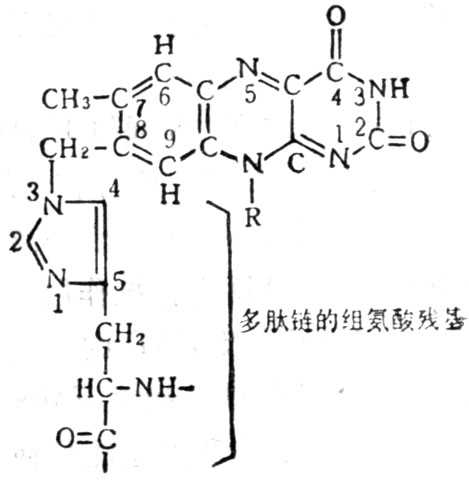
ͼ5-20 ����������ø�е��鰱��
FMN��FAD��ø����ϣ�һ����ͨ�� 8λ�ϵ��Ǽ���ø�����ϵİ����ᡢ��������������ᡣ��ͼ5-20����ά����B2��ø�����ֹ�����ʽͼ5-21��
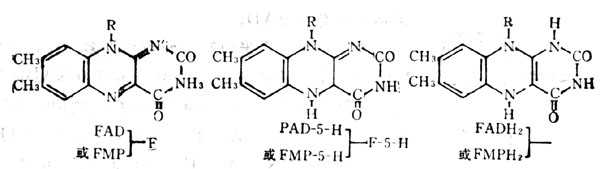
��1����ά����B2Ϊ��ø������ø�ɷ�Ϊ�������ͣ�
�� ���ؽ��ܵ��Ӻ��ֱ������γ�H2O2������������ø��
ͼ5-21 ά����B2��ø���ֹ�����ʽ
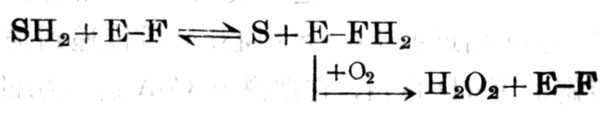
D-������+O2��D-����������+H2O2
�� �γɵ���-ø����ʮ�岢���������γ��м��壬ֱ����O2���ú˵��������븴��ʮ��ֿ�����d,1-����������ø���м������Ϊ��ø�������ż������O2���ú��ǰ�����ˮ��Ϊ��-ͪ�ἰ���������������͵�ø��H202��
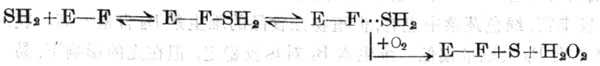
�ۻ���������ø��Ϊ���ӣ��ж�����㣬һ�������1��Moԭ�������Fe-S���ģ�Fe-S����Ϊ��ά��Mo������״̬����һ���ΪFAD���ӡ���������������Mo��6�ۣ����ã�����FAD�����γ�FADH2�γ�FADH2����������������γ�H2O2��
������+H2O2+O2������+H2O2
�ܻ�Ϲ�������ø�����ص�����ø���������ø���������е�һ����ԭ�ӵ�������ȥ��һ��ԭ���γ�ˮ������ﱾ������������ˮ�߽����ڲ���������ø���䷴Ӧʽ���£�����-XH��Ϊ-OH��-NH2��
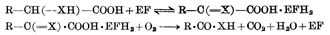
�����NADPH����NADH����Ϊ��ԭ�����γ�ˮ�ߣ������ⲿ��������ø��һЩ��ʮ���廯������Ȼ��������Ȯ�����Ȼ�ø����Ҫ��ø����Ӧʽ���£�
AH+NAD��P��H+H++O2��A-OH+H2O+NAD��P��+��AH���
ʵ������Щ�Ȼ���ø�ĸ�ø���˻��ز��֣�������Ϊ�������ߣ�����NAD��P��H�����ֻ�ԭΪ��ԭ���ߡ�
�������Ϲ��ܰ�����ø����ø�ڸ������У������Ϊ���װ�������Ϊ�װ��������ص����⣬����ҪNADPH�����IJμӣ��˷�Ӧ����-S-S-�Ժϳɵ���˫�����
��2��ά����B2����ø���������ص�������ø�������ж���������������ø��dihydro lipoamide dehydrogenase���������Ļ�ԭø��glutathione reduetase��������������ԭø��thioredoxin reductase����NADH-ϸ��ɫ��b3��ԭø��NADH-cyutochrome b2reductase����NAD��P��H-ϸ��ɫ��P450��ԭø��NAD��P��H-cytoehrome P-450reductase��ǰ����ΪNAD��P��˫������ԭøϵͳ��ͼ5-22����
NADH-ϸ��ɫb3��ԭø��NADH��Hת����FAD���γ�FADH2�ٽ�����ת�Ƶ�������ϸ��ɫ��b3�Ͻ��仹ԭ��NADPH-ϸ��ɫ��P450��ԭø���ӹ���ΪNADPH��������һ����FAD��һ������FMN��������NADPH-ϸ��ɫ��C��ԭø����2������FAD��
ͼ5-22 FAD-˫��������ԭøϵͳ
�������Ļ��ص�������ø�����ܶ࣬�����������к���NAD��P��H������ø�������о��϶���ΪNADH����ø������������ø���������������ữ����ϵ������������ø��FAD��2��Fe-S����ż�������븨øQ������NADH-��øQ����øÿ��FMN��4��Fe-S���ģ�ÿ��������2��4����ԭ�ӣ�NADH��ԭ���ص��ף�Ȼ���Ӵ��ݵ���NAD���Fe-S���ģ��ٴ��ݵ��븨øQ�����Fe-S���ģ���ݵ���øQ�ϡ�
�����������������ܽ��FAD���ף���������CoA����ø�����������븨øQ�Ļ�ԭ��ϵ����������FAD���������Ӵ��ݵ��ף�electron-transferring flavoprotein,ETF����
5.7.4����Դ
����ϸ�����Ժϳ�ά����B2����Ϊ�����࣬��Ҫ��������ʳ���й�����ţʮ��ʮ��������ά����B2�ȽϷḻ����ɫ�߲���Ҳ�У���ֲ��ʮ��ʮʳƷ��ά����B2�������ߡ��ҹ���ֲ��ʮ��ʮʳƷΪ����ά����B2��ȡ��ƫ�͡�ά����B2�Ƚ��ȶ������ڹ��Ӱ���£������ƻ���ţʮ��ʮ�е�ά����B2��¶�ڹ��У��ڲ�����ζ������ǰ����ֽ���ѷ����������ڹ���4h��ά����B2��ʧԼ71%�������ʧ��С�����罫���ˮ����������ˮ��ά����B2���ʧ����ά����B2����ʳ���У��ڴ��ع���Ҳ�Ƚ��ȶ�������ʮ��ʮ�У��䰱�ܷ������£�����1��2�꣬���ܱ���50%���ϡ�
5.7.5����Ҫ��
��1��Ӫ������ָ�� һ����������ά����B2��ʮ��������ϸ��ά����B2��������ϸ��������ԭø������Ϊά����B2��Ӫ��ָ�꣬���н�Ϊ������Ϊ��ϸ�������Ļ�ԭø������μ������ж��������߰�����ø��pyridoxamine 5'-phosphate oxidase���������ȱ��ά����B2����ø�����иı䣬ȱ��4��ʱ�ﵽ����ȱ���̶ȣ���ø����Ҳ�������ֵ�����ָ�����ڶ�������Ρ�
��2����Ҫ�� ���ҹ�ά����B2��Ҫ����������������ÿ����ȡ��0.70��1.16mg�ߣ������ڹ���10�ܣ�����ʮ��ʮͷ������ƽ���⻬����ήʮ������������ȱ��֢����ȡ��12.53mg�飬����ʮ����������������ת�۵�����൱1.2mg��ȡ���������Դ�Ϊ�����Ҫ����1.5mgΪ����Ҫ��������Horwitt�ۺ���������֮�����һ�µġ�����Ϊÿ����ȡ����0.8��0.9mg2��֮�ã�����ȱ��֢��ij��֣�������ʮ������������ȡ��1.2mg,ά����B2����֯�п������������档��ȡ��0.55��1.10mg��d-1����ά����B2��ʮ����Ϊ��ȡ����10%��ÿ����ȡ��1.3mg�����ʱ����ʮ��������20%��
ά����B2����Ҫ�����������Ͷ�ǿ��û�й�ϵ�����뵰������Ҫ���й�ϵ������Ѹ�٣����˻ָ��������벸ʮ��ʮ�ڵ�������Ҫ���ӣ�ά����B2��Ҫ��Ҳ���ӡ���Ӧ��Ӥ��ÿ��0.4��0.6mg,��ͯ0.6��1.2mg����ʮ��1.3��1.7mg���и���ʮ��ʮĸ1.8��2.0mg�����Ƽ���Ϊһ��6mg��һ��2mg��d-1,ÿ��3�Σ�����ά����B2�����ܽ⣬����������Ч����һЩ�����ü���ע��25mgά����B2��Ϊ��Ԥ���ҹ�ά����B2ȱ������ά����B2�¹�����150mg���൱��75mgά����B2�����������ע�䣬�����ͷŻ�����ע��һ�ο�ά�ֳ�ʮ��ά����B2������2��3��֮�á�
5.8�������
5.8.1���ṹ��ʮ��ʮ��
������������ᣬ����������ʮ��ʮ��������Ϊ���������������������nicotinamide�����ṹʽ��ͼ5-23�������Ϊ����ˮ�Ľ��ȶ��İ�ɫ�ᾧ����230��������������ˮ������ˮ����ʮ��ʮ�У�25��ʱ��1g������60mlˮ��80ml��ʮ��ʮ�У������������С����������ױ������������������������ܽ⣬1g������1mlˮ��1.5ml��ʮ��ʮ�У���������Ҳ���ܽ⡣
ͼ5-23 �������������ṹʽ
5.8.2����л
�������ͨ��ATP�����γ�NAD��NADP����������ʮ��������������٣���Ҫ��ʮ�����л����N��-�����������N����-2��ͪ-5-��������ǰ��Լռ�����˵���ʮ������20��30%������Ϊ40��60%��Ѫ���������������������Ϊ0.5��g��ml-1,�졢��ϸ���к����϶࣬ȫѪΪ30��g��ml-1,��ϸ��Ϊ90��g��ml-1,��ϸ��Ϊ70��g��ml-1��
5.8.3����������
�������Ϊ������ø�����ɳɷ֡���ø��Ϊ������������ʶ������ᣨNAD����DPN��������ø��Ϊ������������ʶ����������ᣨNADP����TPN����(ͼ5-24)�����Ƕ�������ø�ĸ�ø����Ĵ���ͨ�����������4λ������һ����ԭ�ӡ����������ַ�ΪA��B������λ��ijЩ�����������ø��������רһʮ��ʮ��ͼ5-25����������רһʮ��ʮ����Ϊ�������棺һ����ֻ��һ�������칹��ĵ����д����ã���ʮ��ʮ������ø����øֻ������ʮ��ʮ�ᡣ��һ��NAD����NAP����ø��ԭʱ����ԭ��ֻ��һ���ض��ķ���ӵ���ऻ��ĵ���λ̼ԭ���ϣ��еļӵ�A�࣬�еļӵ�B�ࡣ
ͼ5-24 ��ø��Ľṹ
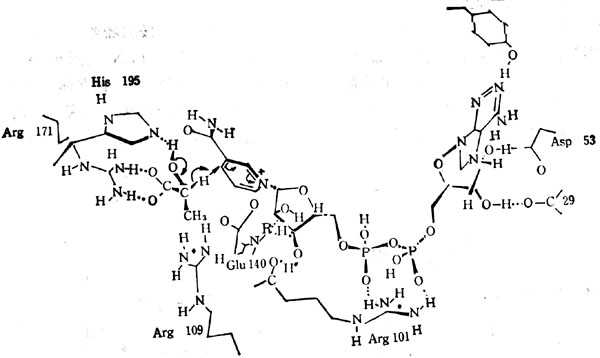
��Ҫ��ø�������ø�������֣���Щ����ø�ӵ�������ȡһ���⣬�������ӣ���ṹͨ��X������ͼ��������ø��NAD��������Ľṹ��ͼ5-26����ʮ��ʮ������ø��NAD��������Ĺ�ϵ������ʮ��ʮ���NAD��һ��H��������ǻ���ø��ʮ��ʮ���ᣨ171�����һ������������鰱�ᣨ195����OH������ȡһ�����ӡ���������ø�Ĺ�����֮��ͬ��

ͼ5-25 �������4λC������H�����幹�죺A��B����λ��
ͼ5-26 ʮ��ʮ������øʮ��ʮ�ἰNAD����ʮʮ��ʮʮ������
��NAD��Ϊ��ø������ø��Ҫ����������ã�������ӵ��ﵽ���ĵ��Ӵ������õ��м价�ڡ�����NADP��Ϊ��ø����ø�࣬��Ҫ���ֽ��л�м����ϵĵ���ת�Ƶ�����ϳɷ�Ӧ������Ҫ���ӵ��м����ϡ�
NAD������NADP��������������������м������ͣ�
��1�� �����ɴ����ȩ��ͪ
��2�� ��ʮ�ʮ����A-��ȩ������
�Ȱ�����ø�ǹȰ����Ѱ��ĵ�һ�����γɦ�-�ǰ�������ᡣ
�Ȱ��� ��-�ǰ���������-ͪ����
��4���ɲ����ͺϼ�������C����C�����������ͼ�����7-����̴�ת��Ϊ���̴���7��-H����NADPH��������8����ӽ��ʶ������ڵ��̴��ϳ��У��̴����嵰���ڰ�Һ���γɵ���ϩת�������廷������ʮëʮ֬�̴�����ת��Ϊ���̴������̴�ת��Ϊ���̴���Ҫ��NADPH����25λ�⣬�ӽ����й���24λH����
���̴�������24-����̴� ���̴�
��5�������תΪ����������ø�������Ӧ����ø��ΪH���Ĺ�������塣���װ��ỹԭø������ΪNADH2+1-�װ����NAD��C+2-�����ᡣ�����Ļ�ԭø������ΪNAD��P������H2���������ġ�NAD��P��+2G��ԭ�������ġ�
�ڦ�-ͪ������øϵͳ�У�����������øͨ��NADPH������ص������ã���Hת�ơ�
��6����������ͨ��������4-������ø�����������Ȼ�ø�������ã�����Ұ��ᡣ���ø����Ϊ������ø����Ϊ������ø���Ϲ��ܼ���ø�Ĵ��������������ǽ��������е�һ����ԭ�Ӽӵ������������γɶ��Ȼ�����һ����ԭ�ӻ�ԭ��ˮ����ԭ����NADPH����Ӧ���������У�
NADPH+H��+����������ʡ�NADPH��+����������ʡ�
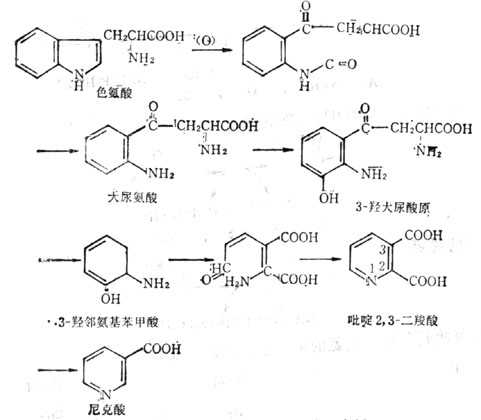
ͼ5-27 ɫ����ת��Ϊ������;��
1-��������+�����������+O2��1-�Ұ���+�����������+H2O��
5.8.4����Դ
�����������������Դ��ʳ�ﺬ�����⣬�п���ɫ����������ת��Ϊ��Ȯ���ᵽȮ���ᣬ����1-Ȯ����ˮ��ø�ֽ�Ȯ����������Ϊ3-�Ȱ��������ᣬȻ����5������ǽ���������£��ɲ�ʮ��ʮ��ε�øϵͳ�������ᡣһ��˵����60mgɫ�����൱��1mg����ᡣʳ���������ĵ���Ϊ����ἰɫ����ת��Ϊ�����֮�ͣ���5-16������ת���������˶��죬�����и�ת������3����������Ů����ʮ��ʮ�ؿɴ̼�ɫ��������ø������ɫ����תΪ���������е���������ø������и����ڷ�ҩ��ת��������ǿ��
��5-16ʳ��������ᵱ��(mg��1000Kj-1)
| ����� | ɫ���� | ����ᵱ�� | |
| ţʮ��ʮ | 5.08 | 283 | |
| ��ʮ��ʮ | 10.3 | 1681 | 32.1 |
| ţ�� | 103.7 | 5376 | 193 |
| ȫ�� | 2.5 | 4830 | 45.4 |
| ������ | 4.8 | 256 | 11 |
| ��� | 10.4 | 1247 | 32 |
| ������ | 7.7 | 294 | 13 |
| ���� | 21 | 445 | 28 |
�������н���͵�����ᣬ�������á��ɷ�Ϊ���ֽ�������������������12��000��13��000����ϣ�����̼ˮ�����ϳɸ���ʮ�壨������Ϊ370������Ƥ�������ֽ���͵�����ᣨniacytin��,�ü���ȡ�����ˮ�⣩����������ⶨֵ����ʮ��ʮ����ʮ��ʮ��ȡҺ�ĸ�20%������ͨ��������Ʒ�ֿ�ʹɫ�������ӣ�Ҳ��ʹ���Ϊ�������Դ���ҹ�����������ɫ����Ʒ�ֵ��������������ȱ���������Ƥ����
��������������ʹ�Ƥ��������Jowar��һ��ʳ�����ἰɫ���������٣���������������Ҳ��ʹ�Ƥ���������ڴ�����ҵ��������У������������ᣬ�������-2-3-���������ӣ�����Ӱ��ɫ������������ת�䡣�����˵�������ʳ�������������������������श����ᣬ���Ļ�����᰷��-6���ͪ����ʮ�������١�
5.8.5����Ҫ������ʮ��ʮӦ��
����ṩӦ��Ϊ27.7mg��100KʮJ-1��Ӥ��0��6����Ϊ33.6 mg��100KʮJ-1,2/3��Դ��ɫ���ᣬһ��75g����������ᵱ��NE15mg��6���¡�����ʱ��Ϊ27.7mg��100KʮJ-1���и�����2mg��ʮ��ʮĸ����4mg��
�����ȱ��֢���������������ơ�����ἰ����᰷�����Ƽ�Ԥ���Ƥ������ͬЧ��������ҩ���ϼ���ʮ��ʮ�϶��߲���ȫ��ͬ�������3g������ҩ�����ã���ʹѪ�е��̴�����-֬�����������������͡��������ڼ��ٴ�֬����֯�ж�Ա����֬���ᣬ�����Ӽ�����ļ��ĸ���ԭ�����á���˳���ʹ�����������Ըβ��������ܵ����鰷���ͷţ���ʹѪ�����ţ�����Ϊ��Ҫ�ĸ����á�����ڸ�ҩǰ15min���Կ��鰷ҩ�������������졣������������Ѫ������ˮƽ��ÿ��3g��������5�꣬����ʹѪ�����ᳬ��8mg%���ڴ����ʱ������������Ᾱ����ʮ��������Ѫ���������ӣ����Դ�����������ȡ������ʮ��ʮʹ��֢��
5.9������B6
5.9.1���ṹ��ʮ��ʮ��
ά����B6��VB6����������ʽ�����ߴ���Pyridoxine,PN��,����ȩ(Pyridoxal��PA��PL)�����߰���Pyridoxamine,PM������������ʽͨ��ø�ɻ���ת����PL��PM���ữ��仯��ø����������ȩ��PLP�����������߰���PMP������ṹʽ��ͼ5-28����
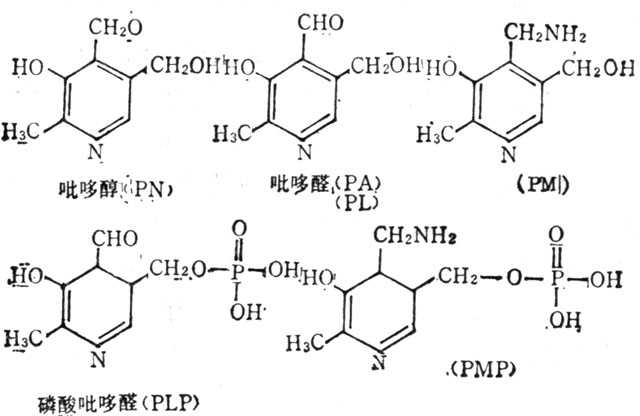
ͼ5-28 ά����B6���丨ø�Ľṹʽ
�ˡ�����С����������������Щ���ﶼ��Ҫά����B6��ţ�������ڳ������ܺϳɣ�����Ҫ��繩�������ߴ�Ϊ�˹��ϳɵIJ�Ʒ����ֲ����Ҳ�У�������Ϊ205.6��ϵ��ɫ��״�ᾧ������ˮ������ʮ��ʮ��Һ���ȶ����ڼ�ʮ��ʮ��Һ���ױ������ƻ����ڶ������ڣ����Ը�øPLP��PMP����ʽ���ڡ���Щ�������4-�������ߴ����п�ά����B6�����á�
5.9.2����л
ά����B6�ĸ���ʽ�����ڵı仯���£�
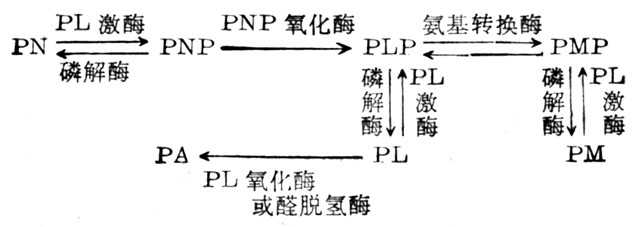
PN��PL��PM�����������PL��ø��ATP��Pyridoxal-5'-phosphotranferase,PL Kinase�������������ڸ���֯����Ρ��Լ���ϸ��������PH����������Ҫ��������������һ����PNP��PMP������ø(Pyridoxic Acid,PA)Ϊ��л���ղ����������ʮ�����ڸ���PL��ȩ����ø���γɣ���������֯������ҪNAD��ȩ�Ѱ�ø�����γɡ�
��1������������ ʳ����ά����B6ΪPLP��PMP��PN��С��ǻʮ�ڱ����ɷ�����ʮ��ʮ��ø��nospecific phosphohYdrolase���ֽ�PLP��PMPΪPL��PM��������ʽΪPL��PM��PN������ʮ��۲��У����輢��������PN��PL��PM���ڸ���PN��0.5��3h�ﵽ��ʮ�壬����С��0.5��4mg��ʱ��Ѫ��ά����B6ˮƽ��3��5h���ָֻ������Ƽ���ʱˮƽ������PL��Ѫ��ά����B6ˮƽ������PA���߽Ͽ죬��PM���մ�л����PN��PLҪ��һЩ��������PLP��������10mg)ʱ��Ѫ��ά����B6��PLP��24h�ڼ���������ά���ڸ�ˮƽ�ϡ�
��2���������л PN������С��ճʮĤ����Ѫ���У�Ҳ���ڳ�ճʮĤ�кϳ�PNP��ԼΪ����30.6%��Ѫ����PN����ɢ�������У�Ȼ�����ữԼռ������10.4%��15.7%������ʮ�����PN��Ѫ��PL��������12����Ѫ����PLP��ռѪ����ά����B6��60%�����뵰�����ϣ�����Ϊ����ϸ�������á�Ѫ����PL�����ϲ��ι̣�Ϊ�������ʽ���ܱ���֯��ȡ�������������ΪPA��PN��PLͨ����ɢ���뵽��ϸ���У���Ϊ��ø���ữ���˵ĺ�ϸ���ɽ�PNP����PLP����������������˹��ܡ�PN�ڳ�����ϸ��PL��ø����Ũ��ʱ������3��5min���뵽��ϸ���У�ϸ���ڵ�Ũ����������Ũ��һ�¡�PL��Ũ�ȳ�����ϸ�����ἤø��Ũ��ʱ�������ϸ���������ӣ�ʹ��Ũ�ȱ���������Ҫ����������PL��Ѫ�쵰�צ�-����ĩ���Ӱ������ϣ�����PL�ں�ϸ���л��ۣ����ں�ϸ���е�Ũ�ȿ�ΪѪ����֮4��5������ϸ���е�PL����Ҳ��һ�����䷽ʽ��
��Ҳ��ά����B6��л��Ծ����֯��PNΪ��ϸ����������ΪPL��ø��PNP����ø���ö�����PLP��Ȼ���پ������ö�ת��ΪPL������ѭ��ϵͳ�У����������ἤø����֯�γ�PLP��
������Ƭ������������У�û��PN���������ữ��������Ϊ����PN��Ѫ���Լ�Һ�ij�������ά����B6ȱ���Ķ������PL��ø�����½�������5�ܵĴ�����У�PL�μ�ø�½�50%�����н��½�14%����Ҳ˵��ά����B6����ϵͳ����Ҫʮ��ʮ��
��3������ ά����B6��Ѫ���п���ɢ�������ж����ữ����PN�������ӣ�������PNռ�����İٷ������ӣ���PNP�İٷ������١��ڼ�����δ����PNP������PLP���ڴ���60%ά����B6�ڼ����У�����75��95%����ԭ���ữø��Glycogenphosphorylase������ϵ����øռ�������ʮ��ʮ����֮5%������Ϊά����B6�Ĵ��泡����ͨ�����⵰��ת������ά����B6�ֽ���������������Ҫ����
��4����ʮ�� ά����B6����Ҫ��л����PA���ɴ���ά����B6��ȡ������20��40%������PAֻ��Ϊ��������ָ�ꡣ�����ܴ������ڵĴ��档���г�PA�⣬����С����PN��PL�ȡ�������������ʱ����3h�ڴ���PA��ʮ����PN����С���л��ۣ���PNŨ�Ƚϴ�ʱ����������ʮ�������PN����10mgʱ������PAռ�����İٷ������ͣ���PN����ʮ�����ࡣPL���ױ����ų���Ҳ���ױ������룬����������ữ��ʽ���ۡ����˿ڷ��������100mg��PL��PM��PN����36h�ڴ�ԭ���������ʮ����
5.9.3����������
����ά����B6�ĸ�ø��Ҫ���л�йأ�����֪��60����ø��Ҫά����B6��������Ҫ���÷������¡�
��1������ת������ ת��ø��ҪΪ�Ȳ�ת��ø��ȱ�ת��ø��ת��ø�ж���PLPΪ��ø��
ת��ø�ڶ����ġ��ݡ�����ʮغʮ�輰���к������ܸߣ������ת��ø��Ҫ��-ͪ�������Ϊ���������塣��ˣ���ͬת��ø����ż������������֮һ������-ͪ������Ȱ�����������ʮ��ʮ�ģ�������һ����������Ѱ��İ����ᣩ���ϸ������ʮ��ʮ��ij��ø��ijЩ�������нϴ�Ĵ����ã���������������Ҳ��һ�����á�ø���������Ǹ��ݾ�����������İ���������ġ����磬�ڶ�����֯��ռ���Ƶ�ת��ø���Ŷ�����ת��ø�������Ŷ��������Ϊ��Ϊ���壬��-ͪ�����Ϊ���塣ת��ø��Ӧ������ġ����������ڣ�ת�����ý��ӽ��а�����������ֽ⣬�ɴٰ�����ת��������һ��������С�
��2�����Ȼ����� �����������γɲ���������ø��רһʮ��ʮ�ܸߣ�һ�ְ���������øֻ��һ��1-�����������á���������ø�У����鰱����ø����Ҫ��ø�⣬����������ø����PLPΪ��ø����Щ�������ö��ڲ�ʮ��ʮ�����֯����Ҫ�����������ʵ��γ��������������á��緼ʮ����1-����������ø�����ҡ��顢��ͣ�ɫ��������ȣ��γ���Ӧ�İ������Ұ����鰷����Ͱ���5-��ɫ���������ǻ������ȱ��ţ���ᡣ�Ȱ�����������ϵͳ�����γɦ�-�������ᡣ
ɫ����Ĵ�л;���ж���һ����ת��ø��Ӱ���·ֱ���Ȯ����ͻ����ᣬ������Ȯ���ỹԭø�����£�����ڰ������ἰ3-���ڰ�������������ᣬ�ٱ������ᡣ�ڴ�л;��������B��ά���ز��뷴Ӧ����ά����B6�й���������ͼ5-29��������ά����B6ȱ��ʱ���Ը��ַ�ӦӰ��̶Ȳ�ͬ���������������ɵķ�Ӧ�ϴԻ���������Ӱ����١�����B6ȱ��ʱ�����������ɣ����л�������ʮ�����ࡣ
��3�������ֽ����� ���ǻ����հ����˿����ɷֽ�Ϊ�ʰ��ἰ��ȩ���ȩ�����˷�Ӧ��øΪ˿����ת�ȼ�ø����ø�ܹ���˿������հ������ְ����ᷢ������ȩ���ѷ�Ӧ����ø��øPLP��
5.9.4����Դ
ά����B6���ڳ�����ϸ���ϳɣ�������������Ҫ������ʳ���зֲ��Ϲ㣬�⡢���ࡢӲ����ˮ�����߲��ж��С�ֲ������PNΪ������������PLP��PMP����Ϊά����B6�Ϸḻ����Դ��ʮ��ʮ��B6�ĺ�����ӳĸ��ά����B6��Ӫ��ˮƽ��ĸ��ÿ����ȡ��2.5mgʱ����ʹʮ��ʮ��ά����B6����Ϊ0.2mg����ȫ����ά����B6,85%�ĺ�������ĥ�ۼӹ�����ʧ������ά����B6ǿ����������У�������Ч��Ҫ��ȫ������߸�һЩ����ȫ������ߴӴ����ʮ���϶ࡣ������ά����B6����Ч�ýϵͣ����������Ҳ����ʧ������Ѫ�ӹ�����������У���������Ч�ü��ͣ�����ֻ��40��50%�������ʮ��ʮ��������������Щ������PLP��ø�������������γɵ�ϣ����ԭ���γɦ�-�����������ᣨ��-pyridoxyllysine����ʹ������Ч�ü��͡�һЩ����ʳƷ��ά����B6����������5-17�С�
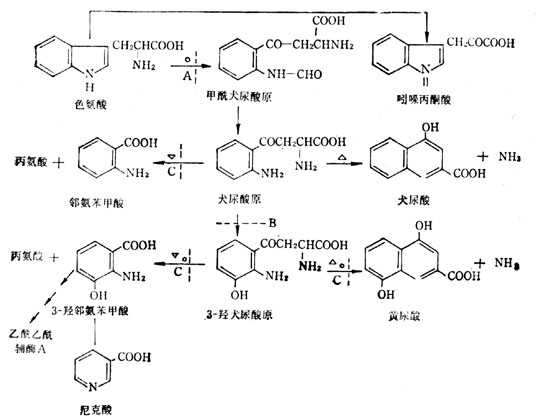
ͼ5-29 ɫ����ķֽ��л
5.9.5����Ҫ��
��1��ȱ��֢�� ά����B6Ϊ�����ᡢ�Ǽ�֬���л�ϵĸ�ø���ٽ��ϳ�һЩ����������Ӱ���5-��ɫ��������ά����B6ȱ�����Ե���������ʮ��������ήʮ����֬���Ρ����ʡ�ƶѪ����ʮֳϵͳ�����ƻ���ˮ����ʮ��ʮ������ҲӰ��ά����B6���и���Ӱ���Ӵ���ϸ���ķ�չ��
Ӥ������ά����B6ȱ�������º�����ֹͣ��������PA��PNҲ�ܵͣ����о���ƶѪ����ɫ����ת��Ϊ���������ɥʧ������ά����B6��ת��Ϊ����������ָ������������������ʧ��
��50������4-�������ߴ���ά����B6�����������ά����B6ȱ�����������ۡ��ǡ��ڲ�����չ����ǰ�üʮëʮ���������Ƥ֬�������ʱ��ʮ�ʮ��ʮ�ʮ�Ҳ�Ҳ�д��������ز���������ʮʪ��λ�в��ã������Ƥ֢��Ƥ��Ҳ�������ھ���ǰ�ۡ��⼰ʮ��ʮ�������ɡ����ס�����֢״�������ȱ��֢״�����ơ�������3����Χ��ʧ���������Ǹй٣�Ȼ���˶�����ʧ������Щ��ʮ��ʮ��״̬����������Į��?��?����ʮ�С���ʳά����B6ȱ�����ߣ���ʮ��ʮ�������������ƸУ���ʮ��ʮͷ��ʮ���ף���ǻƤ֬�����
��һЩ���������У�������Ƥ����ȱ������ȱ������֬���������ƣ�������֮��ϵ�в����ˡ���֬�ſ����Ƴ�ά����B6ȱ��֢��ij��֣��ߵ������ϸ����յ������͵�Ƥ��֢��������
��5-17 һЩʳ����ά����B6������g��100g��ʳ��-1
| ʳ�� | ά����B6 | ʳ�� | ά����B6 | ʳ�� | ά����B6 | |||
| ���ࣺ | ��Σ����� | 840��840 | ���ࣺ | ��� | 40 | �߲ˣ� | �˻� | 210210 |
| ţ���⣨���� | 330330 | ȫ����� | 180 | ��ײ� | 160160 | |||
| ţ�������� | 430430 | �ף�ǿ���� | 170 | ���� | 5555 | |||
| ţ�� | 250 | ��ۣ�ǿ���� | 170 | �㶹 | 160 | |||
| ���� | 350 | �ƶ��ۣ��� ֬�� | 60 | ���� | 250 | |||
| ���� | 650 | ˮ���� | ƻ�� | 724 | ���� | 218 | ||
| ���� | 320 | �㽶 | 30 | �Ϲ� | 82 | |||
| Сţ�� | 340 | �� | 510 | �ܲ�ӧ������ | 100 | |||
| �����⣨���� | 275 | ӣ�� | 54 | �� | 80 | |||
| �㣺 | ���Ŀ�㣨���� | 430 | �� | 44 | �� | 560 | ||
| ���㣨��ͷ�� | 425 | ���� | 34 | ���ף���ͷ�� | 200 | |||
| ���㣨��ͷ�� | 300 | �� | 80 | ���ˣ����� | 150 | |||
| �ݣ� | ���⣨���� | 325 | �� | 60 | ������ | 100 | ||
| ���ⲿ | 683 | �� | 19 | �ɹ��� | �� | 153 | ||
| ������ | ȫ�� | 110 | ���� | 7414 | �Ҹ� | 100 | ||
| ���� | 2 | ��÷ | 55 | ���Ѹ� | 240 | |||
| ���� | 300 | ��� | 86 | |||||
| ţʮ��ʮ�� | ��֬ţʮ��ʮ | 40 | ����� | ���� | 100 | |||
| ţʮ��ʮ | 42 | ���� | 400 | |||||
| ʮ��ʮ�� | 300 | ������ | 730 | |||||
| ��֬ʮ��ʮ�� | 360 | ���ϣ� | ơ�� | 60 | ||||
| ��ʮ��ʮ | 20 | �� | 40 | |||||
��2��Ӫ������ָ��
�ٲⶨ��ϸ������ҪPLP��ø����Ȳ�������ת��ø��EGOT����ȱ�ͪ��ת��ø��EGPT�������ߵĻ�����Ϊǰ�ߵ�1/10�������PLP������PLP֮��ֵ��Ϊָ��������ά����B6Ӫ�������
EGOTָ��=EGOT+PLP/EGOT-PLP
�ڲⶨά����B6�����ͼ��ת�������ø���������PL��ø����ø��PN����ø�����н�PL��ø��ά����B6ȱ��ʱ�������͡�
�۲ⶨѪ����PLPˮƽ�����뼡����PLP֮��ƽ�У��ɴ���ά����B6�����������Ȼ���ڷ��������ķ�չ���ԲⶨѪ��ά����B6�����ڶ�������Ρ�
��ɫ���Ḻ���� ����2��5g1-ɫ�������������л�������ӣ�ά����B6ȱ�������������ʮ�������ӡ�����5gɫ�����6h���л����������25mgΪά����B6����ı��֡�
������PA�ⶨ ����PA��������ʮ��ʮ����졢��ʮ�ˣ���ʮ��ʮ����ʮ��PA֮���൱����������40��50%������Ůʮ��ʮԼռ��������25��35%���ഺ��ǰ��Ů����ʮ����ռ��������2.5��6%��
ά����B6��Ӫ������ָ�����ڱ�5-18��
��3����Ҫ�� ���������Ӧ������ʮ��Ϊ2mg���и���ʮ��ʮĸΪ2.5mg,Ӥ��0.3��0.4mg,�˹���ʮ��ʮ��0.4mg,��ʮŮ��7��9��ʱΪ1.25mg����ʹ����֯�ﵽ���͵ļ�����ͬ������������֯�ﵽ����ʱ�ļ�������ÿ���������к����ƣ�����2.4mg����4.8mg,ʮ��ʮ�ٺ�ţʮ��ʮ9.6mg��
��5-18 ��ʮ��ά����B6Ӫ��״������
| ��Ե״̬ | �ɽ���״�� | |
| ɫ���Ḻ������ | ||
| �������mg��d-1 | >50 | <25 |
| ����Ȯ����mg��d-1 | >50 | <10 |
| ����3-OHȮ����mg��d-1 | >50 | <25 |
| ���-2-3-�������m��d-1 | >50 | <25 |
| ������ʮ���� | <0.5 | >��=0.8 |
| 4-PAa mg��d-1 | ||
| Ѫ�вⶨ | ||
| EGPTָ�� | ��1.25 | >��=1.25 |
| EPGTָ�� | ��1.5 | >��=1.5 |
| Ѫ��ά����B6mg��d-1 | ��25 | >��=50 |
Ӱ��ά����B6��Ҫ�������������м��֣�
����ʳ��ɵ�Ӱ�죺��������ȡ���ߣ�ά����B6��Ҫ��Ҳ���࣬����ȡ100g�����ʣ�ÿ��Ӧ��ȡ1.75��2mg���͵���ʱΪ 1.25��1.50mg��
�ڼ��ص�Ӱ�죺�ڷ���ʮ��ʮ�������ҩ���Ե���15��20%��ҩ�ߵ�ɫ�����л��������˵��ά����B6=����ʹ75%ɫ�����л�������ָ�����������30mg�ߣ�����ȫ��������������������Ӧ��2mgά����B6��Ѫ��PLP����ϸ��ת��ø������������Ϊ��ʮ��ʮ������ɫ�����л�е�Ȯ����ø����״������ά����B6��ø�������ͣ�ά����B6��Ҫ��Ҳ���ࡣ
��ҩ���Ӱ�죺ʹ�������������ν��ʱ��������Χ���ף�����ά����B6����Ԥ������˿���ᣨ�����ҩ������������ϵIJ�����������ά����B6��ʮ�������࣬�����ά����B6����Ԥ����
�ܻ��У�����ʮ����ڣ���Ů��PLPŨ�ȱ�δ������Ҫ�ͣ�ɫ�����л���������ʮ�����࣬Ѫ��PLP�ڻ���һ�ܺ����ӣ��ڷ���ʱ����ˮƽ����ʮ��ʮ����Ȼ�ܵͣ�����ÿ��2��5mg��������֮��
�����䣺������ܵ���ά����B6��л�ϵı仯��ʹ��֯��ά����B6ˮƽƫ�͡�С��������֤������Ķ���PLP������������ӣ�ʹ����PLP���١�
��ʮ��ʮ��Ӱ�죺�Ⱦ���ѪҺ��ά����B6ˮƽ�͡���������֤������ά����B6��ȡ�����٣���ʮ��ʮʹѪ��������PLPˮƽ���͡�
��4����ʮ��ʮӦ�á� ��Seicure������������������ϵͳ�У�PLP���ڹȰ����Ѱ�ø���Ѹ���ø���ײ��ܽ�ϣ�ʹ��-�������ᣨGABA�����٣�GABAΪ������ϵͳ���ʵ����Ƽ������ڸ��Դ����ά����B6��10��25mmg�����������ơ�Ӥ������ĸ��ά����B6��ȡ�����㣬ʮ��ʮ��ά����B6ȱ�٣������˹���ʮ��ʮ���ڼӹ�����������У�ʳƷ��ά����B6�ƻ��������������Ӥ������������2mgά����B6����֮����״��ϸ��Ҳ�����ô����ά����B6���Ρ�ȱ��ʮ��ʮƶѪ֢���ߣ�����2.5mg������ά����B6��������Ѫ�������ټ���ϸ������ʽ��������ʹ֮��ȫ�ָ���������������Ͱ���������ɭ����ͬʱ����ά����B6���ɼ����丱���á������������ƻ����ڵ�Ż�¡����ģ�����Щ���������������������װ������߲���������Լ�����Ĥ�Ļ���ήʮ����gyrate atrophy������ʱ����ά����B6��Ч��
һ��˵��ά����B6��ʮ��ʮ��С��ÿ������1gPN�Թ����������Ӷ���ʮ��Ӱ�졣PM��ʮ��ʮ���һ��ΪPN2�����������ڶ�ʮ��ʮ��С�����ʡ�����������ı����������ô����PN�о����������ʼ�����������?�ĸ������?������֬��ʧ�����˷��ô������ÿ�շ���2��6g������ʱ��Ϊ4��40�£�������ʧ����Զ��֫ʮ���λ�����о�������ʧ���췴����ʧ�ȸо��������ڴ����ά����B6�Ķ�ʮ��ʮ����ֵ�ý�һ���۲졣
5.10��Ҷ��
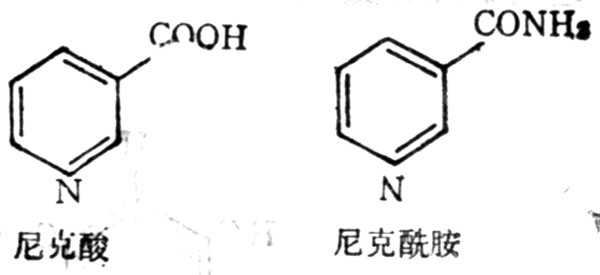
5.10.1���ṹ��ʮ��ʮ��
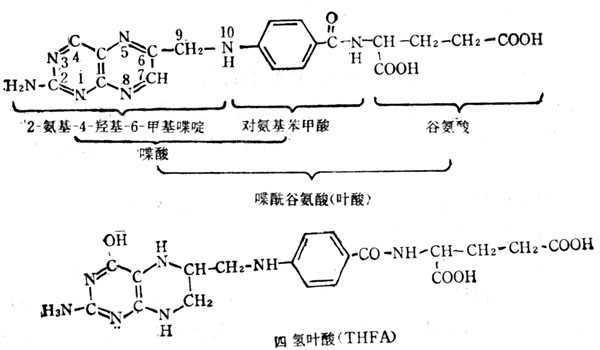
Ҷ����Ƶ����Ȱ��ᣬ���Ľṹʽ��ͼ5-30��Ϊ��ɫ�ᾧ����������ˮ���������κ������ܽ⡣��PH4���±��ֽ�Ϊ��������ण�pteridine�������������ἰ�Ȱ��ᡣ��PH5���ϱȽ��ȶ���������������Ȼ����Ҳ�������ʮ��ʮ����Ϊ���������ʮ��ʮ��Ҷ���Σ�floate����ǰ�塣�Ӳ������˶���ҪҶ���Σ�����-̼������ת�Ƶ��м��塣��Ҷ���α������������㣺�ٽ���ົ���ԭΪ���⼰����Ҷ��������ʽ������Ҷ�ᣨdihydro�� folate,fh2���ڲ�ʮ��ʮ�ද������֯�еĶ���Ҷ�ỹԭø���ã���NADPH�����£���ԭΪ����Ҷ�ᣨteetahydrofolate,THFP�����ڽ��Ȱ������Ŀ���࣬������7��������N5��N10������1-C��ʮʮ��ʮʮ����ЩҶ�����������簱�����ʡ������ʵ��������Ҷ��ԭø����������Ľ�ϣ���ֹ������Ҷ������ɣ��Ӷ�������1-C��ʮʮ��ʮʮ��ת�ơ�
ͼ5-30��Ҷ�������Ҷ��Ľṹʽ
5.10.2����л
ʳ����Ҷ����Ϊ�Ȱ��������ߣ����������ڱ���Ƥϸ����ø���Ϸֽ�ɵ��Ȱ��ᣬ�ٻ�ԭ��THFA���ű�С�����ա���Ѫ����֯Һ����ҪΪN5-��THFA����ϸ�������Զ�Ȱ�����ʽ���档���ڴ���Լ5��10mg������һ�봢���ڸ��ڣ���ͨ��ϸ��������ʱ����Ѫ������֯�н��ø���ֽ⡣Ѫ����������Ҷ���ν�ϵ��ף�һ�ֽ��������ʮ��ʮ��С����һ�ֽ����С����ʮ��ʮ��ǰ��Ϊ���ף�����Ϊ��-�ס�Ѫ���н�ϵ���������Ҷ���Σ�FH2���������ϻ�ԭ���ߣ�THFA��Ҫ�������ڰ������͵����������ڼ��Ի�ԭ��Ѫ����Ҷ���κ�����ʮŮ��ʮ�˶�֮ͯ�������𣬵��и��н���������ӣ�Ѫ��Ҷ���κ����ϸߡ���������Ѫ���������ߣ�Ѫ�弰��ϸ����Ҷ����ˮƽ������ߵġ�
���ڵ�Ҷ������ϸ���е���ϣ�����ϸ��Ĥ�ϣ�����ϸ���ڡ�
��1��ϸ��Ĥ�Ͻ�ϵ��� С����Ƥϸ��Ĥ������Ĥ���Ρ���ϸ��Ĥ�϶������ֵ��ס����н�ϵ���������С��Ҷ���������йء���ǻע��[3H]Ҷ���κ�0.5h������ʮ��ʮ�ں˼�����Ĥ�ϣ����Գ�ʱ������Ҷ������ϸ������������ĵ������ϣ�Ҷ����Ϊ��ԭ�ͼ�����Ȱ��ᣬ�������˵��Ҷ����������Ĥ�ϵĵ������ϣ������뵽ϸ���ڡ�
��������Ե���Ĥ�У�Ҳ�п�����Ҷ���ϵĵ��ס�����N5-��THFA���뵰�����ϡ��ڸ���Ҳ�н�ϵ��ף���Ҫ�ڸε������弰�������֡��������ֵĵ�����ʮʮ��ʮʮ�����Ѿ��ǽ�G-150���������ֳ������֣���FBP-C��FBP-D��FBP-C�������岿ΪMFBP����ϵ�Ҷ��Ϊ��Ȱ����ߵģ�δ�����Ϊ���Ȱ����ߡ�FBP-C���ڸ�����࣬���Ϊ����Ҷ���ι�Ӧ����ʱ���䵰����������δ���֮Ҷ���μ��ٽ϶࣬������ٽ��١�MFBP����һ����DEAE-��ά�ز������ɷֳɶ����֣�һ������2���ʰ�������ø���ϣ���һ�������Ļ��ʰ�������ø���ϡ����Ƿ������Щø�ķ�Ӧ���д���һ��̽�֡�
Ҷ���������м���֭����ʮ������ʮ������������ҪΪ���������������Ȱ��ᣨAcetamidcbenzoyglutamicacid��,��Ҳ�۲쵽ע���ǵ�Ҷ���κַ�������ӵ�֭���ɲ����������ʮ��ʮ��������ʮ������ʮ��ʮ���в����ˡ�
5.10.3����������
THFA��Я1C��ʮʮ��ʮʮ������1C��ʮʮ��ʮʮת�Ƶ��м��
��1��˿������ʰ����ת�䡡˿����ͨ��ת�Ǽ�ø���ã�����̼ԭ��ת����THFA�ϣ�����ʮȥһ����ˮ���γ��Ǽ�THFA���ʰ��ᣬ�����ӦΪ����ģ�Ҳ���ɸʰ���ϳ�˿���ᡣ
��2���鰱���м��л�ϵ����á��鰱���м��л����N-�ǰ����Ȱ�����THFA���ã�����N5-�ǰ���THFA���Ȱ��ᡣ
��3���߰��װ�����N5-��THFA���úϳɵ����ἰTHFA���ڴ˷�Ӧ����Ҫά����B12�μӣ��Ƚ�N5-��THFA�߰��װ���ת��ø�ϣ�Ȼ����ת�Ƶ��߰��װ����Ϻϳɵ����ᣬͬʱ��N5��THFA�����THFA��
��14�����ʵĺϳɡ���5-������ǽ�������ʼ������һϵ��ø�ٷ�Ӧ�������ʺ����ᣬ������Щ������ҪTHFA�����ォ1-C��ʮʮ��ʮʮת�ƣ������ʵ�2��8λC��Դ��THFA��������Я����1-C��ʮʮ��ʮʮ��
��5��������ऺ�����ĺϳɡ�������ऺ���������ऺ����ᣬͨ��������ऺ�����ϳ�ø��thymidytesynthetase�����ã���ά����5��10�Ǽ�THFAΪ������ת������������Ӧ�������ɵ�FH2��ΪFH2��ԭø������NADPH�����⣬��ԭΪTHFA��
Ҷ���ε�ȱ�����¾�ϸ��ʮ��ʮƶѪ��������̬�ϵĸı䣬��������Ҷ����ȱ��Ӱ������л�������Ƕ�������ऺϳɵ�Ӱ�����¡����ٺ�����ΪDNA�ϳɵĹؼ���ϸ������������़�ø����ʹϸ���������γɵ�������ड���һ;������������ϸ���ƻ���ֽ��������������ṩ����ʮ��ʮ����Ϊ������ऺ�����ĺϳ������ơ�������������़��뵽��������ϸ���������У����ڿ��Ժϳ�������ण�������������е���������ɵ�DNA����ϸ��ʮ��ʮƶѪ���˲���������ऺϳ�������ण���ˣ������������е���������व����롣�������˵���������˵Ĺ���ϸ�������뻼�߲�ͬ���Լ�Ҷ����ȱ�������µľ�ϸ��ƶѪ�Ŀ��ܷ���������
5.10.4����Դ
Ҷ��������Ȼ��㷺���ڣ��������ж��С��Ρ�������Ҷ�߲ˡ�����������Ⱥ����ḻ��������Ȼ����Ϊ��Ȱ�������ߡ�������м���¶�ڿ����������ױ��ƻ����ڳ�ʱ�����������ͷ�Ĺ����У����ƻ�50��95%��ţʮ��ʮ���ð���������������ԼΪ168.9nmol/L,�����1min����ʧ2/3������ά����C���ο���Ԥ���ƻ�����������Ҷ��Ļ�ԭ��
5.10.5����Ҫ��
Ҷ���������Ҫ��Ϊ50��g��Ӥ��50��g��1��3��100��g��4��6��200��g��7��10��300��g���и�800��g��FAO/WHO�����Ӧ��Ϊ����ʮ��200��g����ȫ��������£���Ӥ��50��g����ͯ100��g���и�400��g��ʮ��ʮĸ6��8��������ʱ��ϸ����Ҷ��������Ũ����ߣ����Ӥ��û�������������ߡ��ڻ���������ڼ���ʮ��ʮ�ڴ�ĸ��������̥���������࣬ʮ��ʮ��Ҳ��Ҷ���ν�ϵ��ף�������Ҷ���δ�ĸ��ѪҺ��������ʮ��ʮ�У���ʮ��ʮ��Ҷ�����뵰��ϣ�ʹ�䲻��Ϊ����ϸ�������ã�����Ӥ�����ա�������ʮ��ĩ�ڣ�ʮ��ʮĸ��Ҫ�����ӣ������ڶ�ͯ���ഺ��Ҷ����Ҫ�������ӡ�
����Ҷ�ṩӦ���㣬��ʮ��ĩ�ڲ����������о�ϸ����ƶѪ��ijЩҩ�PҶ���������翹��ҩ4-����-N10-��-Ҷ�ᣨmethotrexate,MTX������űҩ2��4-������-5-���ȱ�����-6-�һ���ण�pyrimethamine��,����Ҷ�����������á�������ֹʹ�ô����Ҷ�ᣬ���ó���400��g�����������������ά����B12��������Ϊ�˱������Ҷ������ڸ�ά����B12֮���㣬���Բ������ô�����Ҷ�ᡣ
5.11������B12
5.11.1���ṹ��ʮ��ʮ��
ά����B12��VB12��Ϊ�ܰ��أ�Cobalamin���������ķ�������4����ԭʮ��ʮ������������һ�����ֹ�������������������ᣨ������������������������һ������D-1-����-2-�������������������֮N��ż����ͼ5-31������ѧ�ϳ����ܰ���Ϊά����B12����CN����ܰ����١�CN��Ϊ������ʮʮ��ʮʮ���棬��Ϊ��ͬ���͵��ܰ��ء�һ��ƶԲ�ʮ��ʮ�����������õ�Ϊ�ܰ��أ��ƶԲ�ʮ��ʮ�༰���ﶼ�����õ�Ϊά����B12��������ά����B12���ƴ����
���ܰ�������Ȼ����٣�Ϊ�˹��ϳɲ�Ʒ���ɴ���������ת�����������Ǻ�ɫ�ᾧ�������Ϊ�ؽ�����������ԭ�����ƻ������ڶ��ڸ�ʮѹ������12�棩�����ƻ������ܽ���1��80ˮ�У���ҺΪ��ʮ��ʮ����PH4.5��5ʱ���ȶ���
��øB12����5��-������ǯ�ܰ��أ���ͼ5-32������B12�����ܰ��أ�Ϊ��ʮ��ʮ�ࣨ���ࣩ��֯������Ҫ�ĸ�ø��ʽ��ǰ�����������ڣ������ڰ����ڣ�Ϊ�ϳɵ������������ߡ����ǶԹⲻ�ȶ��������γ�ˮ�ܰ��ء���������±�����ܰ��أ�ά����B12��B12a��B12b��B12c��������ά����B12��ȱ����
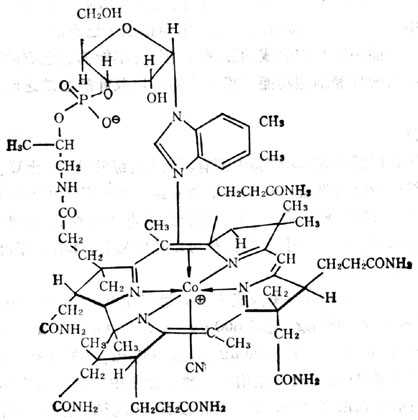
ͼ5-31�����ܰ��ط���ʽ��CN����������ʮʮ��ʮʮ���棩
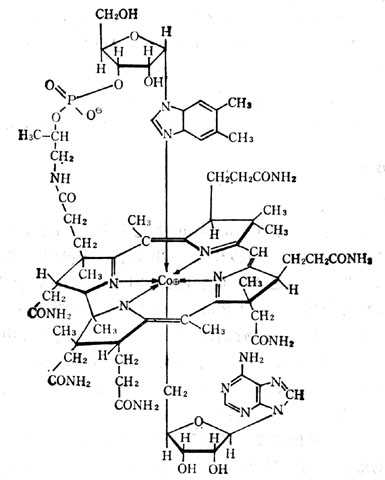
ͼ5-32����øB 12����ʽ
5.11.2����л
ʳ����ά����B12�뵰���ʽ�ϣ���θ����θ����ø�������ͷų���������θ��R����ϣ���PH2��8����£�ά����B12��R��������Ҫ���������ӣ�Intrinsicfactor,IF���������ֱ��50��3������ˣ�ά����B12���ܴ���R�����ת��IF��ϡ���С����ά����B12-R������ֽ⣬��10min��ά����B12ת����IF��ϡ���С����ά����B12��IF��ϡ�R���ײ�����θ���У���Һ����֭��������ʮ��ʮҺ��Ҳ�С�IF��θճʮĤ��ϸ��������ϵͳ����ϵ��ά����B12-IF�Ե���ø���ȶ����м�����ÿgIF�ɽ��30��gά����b 12���볦ϸ���ڣ���ϸ�����������IF�ֿ���ά����B12��ϡ���������ΪIFҲ����ϸ���ڡ�
ά����B12�ӳ�ϸ����ʮ�����ž�����Ѫ���н�ϵ���ϡ���Ѫ����������ά����B12���䵰����transcobalamin��TC��TC��TC��TC��Ѫ��R���ף�Ѫ����80��100%��ά����B12��֮��ϡ�TC���ڸ��кϳɣ�1molTC����1molά����B12�������������ϣ�Ѫ����ֻ��10��20%��ά����B12��֮��ϣ�������Ҫ�����ǽ�ά����B12�ӳ�������������֯����Ρ��������衢��ϸ����̥ʮ�̵ȡ�����Щ����֯���������ܵ㣬��TC��-ά����B12���룬����ø���ڽ���ֽ⣬��������Ϊ�ֽ�ij���Ϊ��ø��Ĥ������ڡ�
TC��TC��Ĺ��ÿ���һ���汣��ά����B12��Ϊϸ�����ã���һ������ܽ�ά����B12��������������ܰ�������ˮ�����������������ȥ������������ǽ�TC��TC��-ά����B12������ĸ���ʮ����뵽��ϸ������ø���ڽ���ֽ⣬Ȼ��ά����B12�������ɸ���ʮ������֭�У��������տ���ʮ��ʮ������IF��ά����B12�������������Ƚ�����
ά����B12�������������䣬��Ҫ���������ٲμӣ���˽�����ά����B12ȱ�����ٴ�ʳ������ȡ�����������䳤����ʳ��ά����B12������ȱ������θ����٣����ֽܷ�ʳ���е���-ά����B12����ʮ�壻��θ�г�̫�࣬û���㹻θճʮĤ�����㹻��IF�����ȵ���ø�������Էֽ�R����-ά����B12����ʮ�壬����IF��ά����B12��ϼ��٣��ݻس�������IF-ά����B12����ʮ���������٣�Ӱ�����գ�����TC�����䵰��ȱ����Ӱ���������䡣
5.11.3����������
1�����˼���ʮ��ʮ�������ж���������Ӧ��Ҫά����B12��Ϊ��ø��
��1��������������������칹����
��һ��ӦΪ����ģ��ɼ�������CoA��λø�����丨øΪ��øB12��ʹ�����ڲ��������С���������-CoA���ɴӱ��ᡢ�������ᡢ�Ӱ�����������������Ϊ������CoA�ɽ��뵽��л;����ά����B12ȱ���ߣ����м���������ʮ�������ࡣ
��2���߰��װ��������ɼ����ᣨ�����ᣩ���߰��װ�����N5-CH3THFA���߰��װ����ת��ø�����£����ܼ���ɵ����ᡣ�����øϵͳ�У���B12Ϊ�������ӣ�����Ҫ���ٸʵ����ἰNADH������5N-��THFAת����ά����B12����ת�����߰��װ����϶���Ϊ�����ᡣ
2��ά����B12ȱ��֢���������ܵĹ�ϵ��ά����B12ȱ��������Ϊ��ϸ����ƶѪ����ϵͳ�ļ�������ϵͳ��֢״�����Ϊ��ʮ��ʮ�ģ�������Χ��ʼ����ָ�д�ʹ�У���չ��������༰���ԣ��������Ҽ��ˣ���������ζ�����������˶�Ҳ�������ȵȡ�����Ҫԭ��Ϊ�������ʡ������������֪����������������ȷ����ϵ��
ά����B12ȱ������һ֢��Ϊ��ϸ���ͷ�Ѫ��ȱ��ά����B12��ƶѪ��ȱ��Ҷ������һ���ġ������á���THFA���塱��˵�����ͣ��ڸ߰��װ���ת��Ϊ������ķ�Ӧ�У�N5-��THFA������ת�Ƴ�ȥ�����THFA�������Ӧ��Ҫά����B12�μӣ����ά����B12ȱ���������Ӧ���ܽ��У�N5-CH3THFA����ת��ΪTHFA������N5-CH3THFAΪ����THFA����Ҫ������ʽ�����Ե���Ҷ�ᣨTHFA������ʮ��ʮ��ȱ���������γ�����THFA��������Я��1-C��ʮʮ��ʮʮ����Ӱ�����һЩ��л�������ʼ�������ऺ�����ĺϳɼ��鰱��Ĵ�л�ȡ�
5.11.4����Դ
ֲ��ʮ��ʮʳƷ��ά����B12�������٣�����������ɺϳ�һЩ���⼰ʮ��ʮ�н϶࣬�˽᳦�е�������Ժϳ�ά����B12�����������գ��ڷ������ʮ������ʳ�߿��ܴ�������Ⱦ��ʳ�Pˮ�л��һЩ��ʳ����ά����B12��ҪΪ��øB12���B12�������뵰�������ϣ������ȶ����ڼ�ʮ��ʮ�в��ȶ�����ά����C������Ҳ���ȶ���ţʮ��ʮ��������������ʧ7%����2��5min��ʧ30%��119��129������13min��ʧ77%��������������143�棩����ʧ10%��
5.11.5����Ҫ��
ά����B12���ڴ�����Ϊ1��10ng������50��90%���ڸ��ڡ���øB12Ϊ��Ҫ������ʽ����B12Ϊ������ʽ�����ڴ����ά����B12����ά�ֲ���ȡά����b 12�ߵĽ�����3��6��֮�á�
�����Ҫ��Ϊά������������������ܵı������Դ��������ά����B12�����Ҫ��Ϊ0.1��g��d-1��
�����Ҫ������λ��d-1��=�����ڿ����õĴ�������/ֹͣ��ȡ����ȱ����ʱ�䣩���գ���
������Ӧ������ʮ�ˡ���ͯ���и��ֱ�Ϊ3��2��4��g������ʳ����ά����B12��50%�����գ���FAO�����Ӧ����ʮ��Ϊ2��g��1��3��0.9��g��4��9��1.5��g��10������2��g���и�3��4��g��ʮ��ʮĸ2.5��g��Ӥ�����˹�ι����0.3��g��
5.12������C
5.12.1���ṹ��ʮ��ʮ��
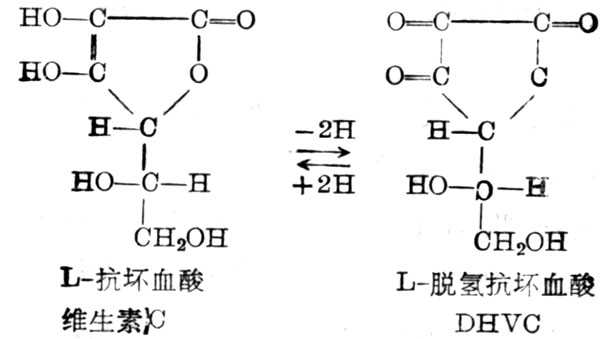
ά����C��������Ѫ�ᣨAscorbic��acid,VC��,Ϊˮ��ʮ��ʮ�ģ����������Ҵ���������֬�ܼ��С�ά����C�ڿ���Ѫ�����������⣬ת��Ϊ���⿹��Ѫ�ᣨDHVC����ͼ5-33����
ͼ5-33��ά����C�Ĺ���ʽ����������ԭ
���ද���ܺϳ�ά����C���ˡ��P�����ܺϳɣ���Ҫ�������ȡ�������������������ȡ���¶�ڿ����У���ʮ��ʮ��Һ����������Cu2����Fe3��,���ܼ�����������ά����C����ΪDHVC����һ��ӦΪ����ģ�����������γ�������ԭϵͳ��Ȼ������DHVC��������ͪ��������(dikotogulonicacicd),����ȡ�
5.12.2����л
ά����C�����ڵı�����������йأ������йأ�������С�������ڱ��������ߣ�������ȫ��������ʮ������ȡ100mg,������ȫ���������ڡ�180mgʱ����76%��1.5g,����50%�������ʱδ�������߲����ڳ��ڣ�Ӱ����ѹ��������к��ά����C�ѱ�����ע��1gά����C���ռ�24h�ⶨ�������˥��Ϊ337h��24h����ʮ��������83%������84%Ϊά����C������Ϊά����C�����������ᡣ����������£�ά����C���Ϊ1500mg�����С������ʮ���С���仯���ֲ�������ˮ��Һ���֣�����ʮ��ʮ�١��ȡ�Ƣ����Һ��ʮغʮ���Ũ����ߡ�ά����Cȱ��ʱ��Ũ���½�������ÿ�ո���25��250mgά����C����ʮ��ʮ�ټ����´���Ũ��������������֯Ũ�Ȳ��䣬��Щ����Ҳ��Ϊ��֯������ά����C����ʮ��ʮ�ټ����´���Ũ��������������֯Ũ�Ȳ��䣬��Щ����Ҳ��Ϊ����֯������ά����CŨ�ȼ�����������������Ũ��Ϊ��ߡ�
��������ά����C�������ⱥ�ͣ������е���ʮ������Ѫ��ά����CŨ�ȵ�ʱ����С�ܿɽ������գ�Ѫ��Ũ�����ߵ�1.4mg%����С�ܲ������գ��Է�ֹѪ��Ũ������������������ʮ����Ginter��Ϊ������⣬ӦΪ������֯�ﵽ����״̬��ÿ��300��600mg�����ɴﵽ����ˮƽ������Ϊ5g��������Ҫ��ʮ�����ά����C��DHVC�⣬����ά����Cͨ��Ѫ�����ϰ��йأ�����ά����C�ܲ�Ѹ��ͨ��������ά����C����ʮ��;����ҪΪ�����ά����C��ͨ������������CO2��ʽ����ʮ�������١�����������ʮ����Ҳ�ɴӳ���������������ʮ����˵��������Խ�ά����C��ȫ�����ֽ⡣
5.12.3����������
��1��������ԭ���á����������ڣ�ά����C/DHVC��������ԭ��Ӧ���ϻ�������SH��/˫���������S����S��ϵͳ����ϵ�����ά����C��ʹ���ڡ���SHˮƽ��ߡ�ά����C�����ڿ����������ʮ�ɻ������ã���Щ��ѧ���ʶԻ���������漰����ʮ�ɻ������ã�������������������������ʮ��ʮ�����Ȼ�̼������ҩ�еİ���ù�ض���������ˡ�ά����C��Ϊ����ˮ��ʮ��ʮ�Ŀ�������������֬��ʮ��ʮ����������Эͬ���ã������ڷ�ֹ֬���������������һ�������á����۾����ڹ�������£�Ҳ�ɲ���O2����ʮ�ɻ�������Ϊ�����ϲ�����ԭ����Щ��ʮ�ɻ������������Ϊ���ڿ���������ά����C������������������Դ�����ά����C������ֹ���ֹ��������õ��ƻ���
��2��ά����C�Խ�ʮ��ʮ��ʮ֯��Ӱ�졡ά����C�Խ�ʮ��ʮ��ʮ֯�н�ԭ������������ʮ��ʮճ���ǵĺϳɶ���Ӱ�졣
��ά����C�뽺ԭ���ϳɵĹ�ϵ������άϸ���ϳɵ������У����ָ����ἰ��������ֱ������Ȼ�ø���������Ȼ�ø������ת��Ϊ�ȸ����ἰ�������ᣬ����ø��Ҫ��-ͪ����ᡢ����Fe2�������صIJμӣ�ά����C�����뷴Ӧ������ʹøFe2������ʮ���е�������2�ۡ�ά����Cȱ���Կ��������Ľ�ԭ��֯�����˿������ϵ������Ե�Ӱ�졣
��ά����C�뽺ԭ���ϳɵĹ�ϵ��ά����C����������������ԭ������á�ά����C����Ϊ�ֽ��������뵰���ʽ�ϵĴ���ӣ��������Ƥ������ʮ��ʮ��ά����CҲ��Ӱ������ʮ�������ٵ��γɣ�����ι�Ե��̴����Ϻ�ע��ά����C��������������������ؼ�����ʮ������-4������ע��������ˮ��Ҫ�ߣ�ά����C�������������������ữ�йء�
��3��ά����C��ijЩҩ���л�Ĺ�ϵ
��ά����C�Ը�����øϵͳ��Ӱ�죻һЩ֬��ʮ��ʮҩ���ڸ�����øϵͳ�����£������Ȼ�����ȥ�Ļ�������Ϊ��ʮ��ʮ����������ڴӵ�֭��������ʮ���ⶾ���л���ɱ�����������ø����������øϵͳ��ά����Ӫ�������Ӧ�Ƚ�����ά����Cȱ���ij̶���δ�ﵽ�����½�ʱ���������ǻ���øϵͳ�Ļ������½���������ϸ��ɫ�ػ�ԭø�ļ�����࣬Լ��85%������ά����C����֯��ά����C���ڽ϶�ʱ���ڻָ���������ø�����ָ�������
�ڶ��鰷�����ã�ά����CӰ���鰷�ķֽ��л�������ȥ�鰷�����á���ι��������ά����C����ʱ����֯��ά����Cˮƽ���Ͷ��鰷ˮƽ������ȱ��ά����C10��12�գ��鰷ˮƽ�ﵽ��ʮ�壬��ά�ִ�������������һ��ά����C5mg��100g����-1�ļ�������ʹ�鰷�½�������ˮƽ����һЩӦ��ʮ�����[�����ij��ҩ����أ��䣨6��1�棩���ȣ�39��1�棩���������м����˵�]����������ά����C��Ѫ���鰷������ԼΪ110��160ng��ml-1��,������ά����C5mg��100g����-1���鰷ˮƽ��������70ng/ml�����鰷��һ��Ѫ���������ã����������Ѫ�ܵ������á�
�۷�ֹ�������������������ε��°����ã��ϳ�4-������������2-�����Ĺ�������ʮ�װ������������Щ���ʵĴ�л��������Ȼ��ǰ����Ļ���������п���Ϊ��ʮ���°����ʣ�ʳ����ɫ����ijЩ��л�����ֲ��С���ʮ���У�Ҳ������������Щ��л�����������������°����ʣ�����ͬʱ��һ����ά����C����ʱ��������ֹ�����������罫�Ȼ��±����ı�����Һ��ʮ��ʮ2h����Ѹ������������20mg%ά����Cһ��ʮ��ʮ�����Է�ֹ����������ÿ��ÿ����ȡ��300mgʱ������ά����C����Ϊ4����5mg%���ɷ�ֹ��Щ��л��������
������֪���������Զ������°����ã��ӹ�������������������ʹ��ɫ�죬ʳ���е������ξ����������ÿɲ����������Ρ��������ο������ٻ��尷���ã���θ���γ��������������������°����á���ʳ�����Ӽ���ɱ���г�������Щ���ࡣά����C���밷�������������������ã������ֹ�������IJ���������������������ͬʱ������θ�У���Ũ��Ψ�������ƣ�ӦΪ�������ε�2����
���뷼ʮ���就�����л�Ĺ�ϵ���Ұ���Ϊ�DZ���İ����ᣬ�����ɱ��������ǻ�ת�����������Ȼ���������Ҫ�����लμӣ���������ʧȥ���ã�ά����C��NADPH���仹ԭ���ָ������á�
�������ڷֽ��л�У��Ⱦ�����ת��ø���ñ�ɶ��Ȼ���ͪ�ᣬ�ٱ�Ϊ����ᣬ��Щ���������Ȼ������裬��Ҫ��-�Ȼ���ͪ������ø���������أ���O2����-ͪ����ᡢFe2����ά����CΪά��Fe2��ԭΪ״̬���Ұ������٣�ά����Cȱ��ʱҲ���Խ��зֽ��л�����Ұ������ʱ������Ҫά����C��
����һЩ�����������õĶ���Ӱ������Ұ���ת��������Ұ����Ȼ�����Ϊ��ͣ�Ȼ��ȥ�Ȼ���ɶ�Ͱ������Ȼ�Ϊȥ����ʮ��ʮ���أ��ټ����仯��ʮ��ʮ���ء�ά����CӪ��״������Щ�Ȼ��������йء�ά����C��ȡ������ʱ�����Ұ����Ȼ�ø����ʮ��ʮ�١��Ρ������ߣ��Ļ���ԼΪȱ����֯��2��4����������������ά����C���ܴٽ��������ά����C�ٽ��Ұ��ἰɫ������Ȼ���������ٽ����������Ȼ��������û�����һ���ġ����ǰ����Ȼ��������У����������ऻ�ԭ�������ड���Ͱ�ת���ȥ����ʮ��ʮ������Ҫ�Ķ��-��-�Ȼ�ø������ά����C����ʮ��ʮ����Ϊ�������ء����-���Ȼ�ø��һ����ͭ���ף�ͭ����ΪCu2���������Ȼ���������Cu2����Cu2����ά����C������ԭ��Cu2������
��ά����C�Ի������գ�cAMP�������ã�ά����C��ʹcAMPֵ���ߣ��������������������ø�����ã�ά����C��cAMP�����ƵĽṹ�����߿ɾ����������ø�ϵ����õ㡣���������ռ��λ���Ѵﱥ�ͣ����ྺ������ά����CѪ��Ũ�ȴﵽ10-4mol���൱��2mg%��������ֹ��ø��cAMP�ķֽ����á�cAMP��cAMP�ṹҲ���ƣ�Ҳ�о������ã�����ά����C����ʱ��ռ����һ����������������õ㣬����cAMP��cAMPҲ���������������á�
��4��ά����C����Ѫ��ϵͳ�����á��ļ�ά����C�������٣���Ѫ�ܷḻ��ά����C���ɰ�ϸ���������ļ��������ˣ��ڷ�����12h�ڣ���ϸ����ά����C������ˮƽ�������ע��ά����C�ɽ��ͼ�ʮ��ʮ��ɽ���˵������ʡ��������ļ��������ˣ�40�ˣ��ֳ����飬һ�鲻����ά����C���ڶ��鲹��1gά����C�������鲹��2gά����C�������ڼƹ���6���¡��ڶ���Ѫ��ά����C��������Ѫ��ά�����ܽ�������Ѫ��֬���ı䡣������Ѫ֬���ı䡣������Ѫ��ά����C����96%��Ѫά�����ܽ���������45%��ѪС��ۼ�ָ���½�27%�����̴��½�12%����-֬���ף�LDL���½���HDL��������ֹͣ��Ӧ���ָֻ�����ǰ��ˮƽ��������һЩ���߶��۲쵽����ά����C��Ӧ���㣬����������أ����������������ද��������������Ӳ�����䡣��Щ�����۲쵽��Щȱ�����Ѫ���μ�Ƥ���ڵ��̴��ĺ�������������ά����C������Ӧ�ߣ����Ը�֬�����̴������ϵ��������֯�У����̴��ĺ���Ҳ��ά����Cȱ�����࣬���������ά����C�ߣ�ÿ��50mg���ϵ͡���ˣ���Ϊά����CӰ�쵨�̴���7λ�Ȼ�����ʹ��ת��Ϊ������������٣�������ֹ���̴��ֽ⼰��ʮ������ʹ�������ڻ��ۡ�����Щ���ױ���ʹ�ô����ά����C���ܽ���Ѫ���̴���
5.12.4����Դ
����ֲ����ά����C�϶࣬����ֲ���е��л��ἰ���������������Ա����������ƻ���������봢�������������ʧ������������ά����C�����϶࣬����4���£���ʣ4���£���ʣ1/2������Ҷ�߲˸�����ʧ�����˴���2�պ���ʧ2/3���й�������������䱣������50��70%��������������Ƚ��ȶ���6λ����������ڿ���֬������ά����C��������Ϊˮ��ʮ��ʮ�ߣ�����ʮ��ʮ����ʮ��ʮ��Һ���ȶ�������Cu2����Fe2�����������ƻ��������ں濾������Ҳ�����ƻ�������ʮ��ʮ��������ά����C��ͬ������Ļ�ʮ��ʮ������Ϊǿ��ʳƷ�á�
ά����C��ҩƬ��ʽ�����Ч���ȴ���ʳ����ȡ�ߵ�Ч��Ҫ��һЩ����֯��ά����CŨ��ҪСһЩ��������֯��ά����C��ȡ�����ޣ���η��õ�Ч����һ�οڷ�ͬ��������Ч��Ҫ�á�
5.12.5����Ҫ����������ĸ�����
����Ӣ��������ά����C�����Ҫ��Ϊÿ��10mg����Ԥ������Ѫ���ķ����������ڹ�Ӧ�����������ϴ�������Ϊ���Ϊ1500mg��ÿ��ת����Ϊ3%����Ӧ��ӦΪ45mg���ҹ�����56��ά����CӪ�������ʮ���������10�ܲ�ͬά����C�����IJ��䣬�ⶨѪ������ϸ������ά����CŨ�ȡ�����������ָ�꣬��Ӧ��Ϊ70mg�����Ա�֤����ά����CӪ���Ƚϳ�ԣ��Ѫ��ά����CԼΪ0.8mg%����ϸ��ά����C28mg���ϣ�����ʮ�������ߣ������ܳ�����ã�ʮ��ʮ�ѽ��١�
����������ά����C��һ�����������ܣ���ʮ��ʮ����ʱʹ�ô����ά����C����ʮ����ú����ò���������Ϊ����Ϊά����C��л�IJ����������ʮ��������������ɲ���ƶ��γ����ʯ����ά����C��ȡ������4gʱ�����в������������ӡ��˻�����ά����C��ʮ��������ʱ��ʹ�����ʮ��ʮ���ӣ��������ܽ�����γ�����ֽ�ʯ��Ҳ����ʹ�硣����������ʱδ�����ᣬ�в���˵��θ�е�������ڸ����͵�ά����B12�У�ֻ��ˮ�ܰ��ؿɱ�ά����C�ƻ����������������ں������࣬����ά����C��ά����B12Ӱ�죬�����ڲ��ܴ���ijЩ����ʱ��ˮ�ܰ���Ϊ�������Ҫ��ʽ��ά����C���ܶ�ά����B12�в�����Ӱ�졣����Ը�������ʱ�������ά����CʹѪ������DHVC�����������ӣ��ɵ��¶�������������ע��DHVC��ʹ�ȵ��ϵĦ�-ϸ���˻���Ѫ�����ߡ��Ը߹����ŵ��ˣ���������ά����C��Ҳ��ʹѪ��DHVC���ӡ���ʳ���Ե����ʴ���һ������ʱ����15%�ҵ����ţ��������������Գ���ͨ��ʳʱ���˸����á�
�����ά����C�������ӻ����ά����E����Ҫ�����˷���5��10gά����Cʱ���������գ��ɱ���ʮ�������¶���Ż�¡�����Ȼͣ������������ڴ�л��ͣ���ڸ�ˮƽ���Ͽ�ؽ��������ù⡣��ĸ���ڻ��м���ʮ��ʮ�ڶ���������ά����C�������ʮ��ʮ��ι��ά����C���ϣ�ά����Cȱ��֢״�������ij���ʱ���һ������Ҫ��һЩ��������ͣ��ά����C����ͼ���ʱ��Ӧ���ؼ��٣�ʹ��������Ӧ���̡�
5.13��������������
B��ά���������з��ἰ�����أ����Ǿ��к���Ҫ���������ܣ���������Դ�㷺����ʮ��δ�����е��͵�ȱ��������
5.13.1������
�����DZ�������ļ��������-����-��-��-�����������϶��ɣ����Ǹ�øA��CoA�����������嵰�ף�acyl��carrier��protein,ACP������ɲ��֡������ڵķ��ᣬ������������ɸ�øA��ACP�ĸ������乹��ʽ��ͼ5-34��
ͼ5-34���������ʽ
��øA�����ǡ�֬�༰�����ʵĴ�л�����Ǵ�л�б�ͪ��ת��Ϊ������øA���ɴ˿ɺϳ�֬�ᣬ������������γ����������������ѭ����12�ְ����ᣨ�����ʡ�˿���ա����ס������������ҡ�����ɫ���ռ��������ᣩ��̼���ֽ��л���γ�������øA��
֬����ֽ���ϳɶ���Ҫ��øA�μӣ���֬����ϳ�ʱ��ACP������֬����ϳ�����Ҫ��ø�γ�һ��������ʮ�塣�����ϵ��ACPΪ���ģ��䳤�����᷺���ϻ��Ұ�������ٲȻ��һ�ڱۣ���һ��ø3������֬����ת�˵���һ��ø�����ϡ�
5.13.2��������
��Ȼ����������Ц����¶��֣���������������ͬ���ڳ�ʮ��ʮ���ȶ������ڸ�ʮ��ʮ�������������£���ʹ��ʧȥ��ʮ��ʮ�����Ļ�ѧ�ṹ��ͼ5-35�������ز������Ȼ���ø��������Ħ�-NH2���ϣ���Ϊ�Ȼ�ת��ø������ø�ĸ�ø��ͼ5-36����
ͼ5-35��������
ͼ5-36��������-ø���Ȼ�������-ø�����Ȼ�ת������

���Ȼ��Ļ�ʮʮ��ʮʮ��������Ȼ��Ļ�ʮʮ��ʮʮ��������һ�Ȼ����ڡ�
��Щ�Ȼ�ø�ڲ�ʮ��ʮ�����л��������Ҫʮ��ʮ�����磬��֬����ϳɵIJ����У�����-CoAת��Ϊ������-CoA��Ҫ��������A�Ȼ�ø��ijЩ�������л�����ɱ��ᣬͨ������-CoA�Ȼ�ø���¶����������ᣬ�ٽ��зֽ��л����ͪ���Ȼ�ø�������γɲ����������������ѭ�������һ��ɺϳ��Ŷ����ἰ�γ�����ϩ����ͪ���Խ�����ԭ������
�Ȼ�ת��ø������ø���������£�

��������Դ�㷺������ϸ��Ҳ�ܺϳɣ�������ಢδ����ȱ��֢���ѱ����ʵ����������ȱ������Ϊ�շ����������������Ľ�����ڵ����к���һ�ּ�ʮ��ʮ�����ʣ���Ϊ�������ص��ף����������ϳ�Ϊһ�ַdz��ȶ�������ʮ��ʮ�������յĻ�������徭���Ⱥ����ֿ����ص��ױ��ƻ��������������ؽ�ϡ�
�����
1.Simpson��K.L,Chichester��CO:Ann��Review��Nutr.1:351,1981
2.����??�����ɷ��ȣ��ҹ�����ά����A��Ҫ�����о�����Ӫ��ר��֮��31������ҽѧ��ѧԺ���������о������1980
3��Henry��HL,Norman��AW:Ann��Rev��Nut.4:493,1984
4.Machilin��LJ,Brin��M:Vitamin��Ein;Nutrition��and the��Adult��Micronutrients245,Edi.R.B.Alfin-Slater,D.Kritchevsky��Plenum��Press��N.Y.U.S.A.1980
5.Olson��REAnn,Rev.Nutr,4;281,1984
6.OlsonRE��Ann,Rev.Nutr,4;281,1984
7.Neal��RA,SauberlichH��E:Thiamin��In:ModernNutrition��in��Health��andDisease1916th��ed.Edi.R.S.��GoodhartM.E.Shils��Lea&Febiger��PhiladelphiaU.S.A.1980
8.FlodinN��W:VitaminB6��In:Vitamin/Trace/Proteins��Interaction��Vo14��135Eden��Press��Ine.Quebec��Canada��1981
9.FlodinN��W:VitaminB6��In:Vitamin/Trace��Mineral/ProteinInteraction��Vo14��135Eden��Press��st,Alb ans��Vt.U.S.A.1981
10.Ink��SL,HendersonLM:Ann��Rev,Nutr,4:455,1984
11.Wagner��C:Ann,Rev,Nutr.2;229,1982
12.Goulian��M��et��al:Ncad.Sci.U.S.A.77:1956,1980
13.Sutharum��B,Alpers:DH:Ann.Rev,.Nutr.2:243,1982
14.Flodin��NW:Vitamin��CIn:��Vitamin/TraceMineral/Protein��Interaction��Vol.1488,EdenPrexx��St��Albans��Vt.U.S.A1981
15.�ܵ��ڵȣ�Ӫ��ѧ����4��99��1982
16���չ�ȫ�ȣ��л�Ԥ��ҽѧ��־15��30��1981
17���ܵ��ڵȣ����������о����ϻ�ࣨ����Ӫ��ר��֮����52��1980���